Volume 10, Number 10—October 2004
Dispatch
Novel Recombinant Sapovirus
Abstract
We determined the complete genome sequences of two sapovirus strains isolated in Thailand and Japan. One of these strains represented a novel, naturally occurring recombinant sapovirus. Evidence suggested the recombination site was at the polymerase-capsid junction within open reading frame one.
The positive-sense polyadenylated single-stranded RNA virus family Caliciviridae contains four genera, Norovirus, Sapovirus, Lagovirus, and Vesivirus (1). Human norovirus is the most important cause of outbreaks of gastroenteritis in the United States and infects all age groups (2). Human sapovirus is also a causative agent of gastroenteritis but is more frequent in young children than in adults (3). Most animal caliciviruses are grouped within the other two genera. In 1999, Jiang et al. (4) identified the first naturally occurring human recombinant norovirus, and several other strains were later described as recombinants (5–8). Evidence suggested that the recombination event occurred at the junction of open reading frames one and two (ORF1 and ORF2), but this finding was not proven. Norovirus ORF1 encodes nonstructural proteins, including the RNA-dependent RNA polymerase, ORF2 encodes the capsid protein, and ORF3 encodes a small capsid protein (1). Nucleotide sequence of the polymerase and capsid junction generally is conserved among the human norovirus genotypes (4,6), which likely facilitates a recombination event when nucleic acid sequences of parental strains come into physical contact in infected cells, e.g., during copy choice recombination (9).
We used genetic analysis to investigate a novel, naturally occurring recombinant sapovirus. Two strains were used for the analysis, Mc10 strain (GenBank accession no. AY237420), isolated from an infant hospitalized with acute gastroenteritis in Chiang Mai, Thailand, in 2000 (5), and C12 strain (AY603425), isolated from an infant with gastroenteritis in Saki, Japan, in 2001 (unpub. data). Although the original polymerase chain reaction (PCR) primer sets that detected these two strains were different, both were directed toward the conserved 5′ end of the capsid gene and have been shown to detect a broad range of sapovirus sequences in genogroup I (GI) and GII (5,10). For Mc10, primers SV5317 and SV5749 were used; for C12, primers SV-F11 and SV-R1 were used.
Figure 1

Figure 1. A) The genomic organization of Mc10 and C12 strains. B) the SimPlot analysis of Mc10 and C12. Mc10 genome sequence was compared to C12 by using a window size of 100...
The complete genomes for Mc10 and C12 were determined as previously described (6). As shown in Figure 1 A, the sapovirus genome has an organization slightly different from that of the norovirus genome. ORF1 encodes nonstructural proteins, polymerase, and the capsid protein, and ORF2 encodes a small protein (1).
Figure 2
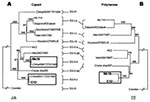
Figure 2. Phylogenetic analysis of (A) capsid (376 nt) and (B) polymerase (289 nt) sequences of Mc10, C12, and additional strains in GenBank. Sapovirus capsid sequences were classified on the basis of the...
Initially, we grouped Mc10 and C12 into two distinct GII clusters (i.e., genotypes), on the basis of their capsid sequences (Figure 2 A) and the phylogenetic classification scheme of Okada et al. (10). In addition, the overall genomic nucleotide similarity between Mc10 and C12 was 84.3%, while ORF1 and ORF2 shared 85.5% and 73.3% nucleotide identity, respectively. These results corresponded with the capsid-based grouping shown in Figure 2 A. By comparing sequence similarity across the length of the genomes with SimPlot with a window size of 100 (11), we discovered a potential recombination site, where the similarity analysis showed a sudden drop in nucleotide identity after the polymerase region (Figure 1 B). Nucleotide sequence analysis of ORF1 with and without the capsid sequence indicated 71.3% and 90.1% nucleotide identity, respectively (Figure 1 A). To additionally illustrate the nucleotide identities of ORF1 minus the capsid sequence, a phylogenetic tree of polymerase sequences of Mc10, C12, and other available strains was developed (Figure 2 B). However, for three strains (Mex14917/00, Mex340/1990, and Cruise ship/00), the polymerase and capsid sequences of ORF1 were not continuous, i.e., they may represent two different strains. Nevertheless, Mc10 and C12 were in the same cluster by polymerase-based grouping but were in distinct clusters by capsid-based grouping (Figure 2). All other strains maintained clusters by polymerase- and capsid-based groupings.
These findings showed Mc10 and C12 had high sequence identity up to the beginning of the capsid region where the sequence identity was considerably lower. These results are easily explained by a recombination event, a single point recombination event occurring at the polymerase-capsid junction. At the end of the polymerase region, there were 44 nt, which included the first 8 nt of the capsid gene and showed 100% homology. After these nucleotides, the identity decreased and was clearly different, as shown in Figure 1 B. This conserved region may represent the break and rejoin site for Mc10 and C12 during viral replication, although direct evidence for this event is lacking.
A sudden drop was indicated, followed by a rise in nucleotide identity between nt 6,250 and 6,500 (Figure 1 B). Although our initial hypothesis was that another recombination event occurred, closer inspection indicated that this region corresponded to amino acids 358 and 440 for the capsid protein and likely represented the hypervariable region, as described recently in the structural analysis of sapovirus capsid protein (12). For norovirus recombinant strains, we also observed a sudden decrease in nucleotide identity in the related capsid region (13), which represents the outermost protruding domain (P2) and is subject to immune pressure (14). For these reasons, a low homology, even between closely related strains, is generally seen in this region (6), although further studies by sequence analysis with other strains are needed.
In a recent study, we genetically and antigenically analyzed two recombinant norovirus strains (13). When the polymerase-based grouping was performed, these two strains clustered together; when capsid-based grouping was performed, these two strains belonged in two distinct genotypes. When we compared the cross-reactivity of these two virus-like particles (VLPs) and hyperimmune sera against the VLPs, we found distinct antigenic types for the VLPs, although a considerable level of cross-reactivity was found between them. We recently expressed C12 capsid protein that resulted in the formation of VLPs, but we were unsuccessful in expressing Mc10 VLPs (G.S. Hansman, unpub. data); therefore the antigenicity of these two strains remains unknown.
Jiang et al. (4) reported two potential parental norovirus strains that were cocirculating in the same geographic region (Mendoza and Argentina in 1995), which provides some evidence for where and when the recombination event may have occurred. In addition, Jiang identified the progeny strain from the event, the Arg320 strain. In our study, Mc10 and C12 were isolated from Thailand and Japan, respectively, but we have no evidence for the place and time of the event. While the genetic analysis for Mc10 and C12 identified a possible recombinant sapovirus strain, the analysis does not clarify which of the two strains was the parent strain and which was the progeny strain. Further extensive studies are needed that perform sequence analysis of polymerase and capsid genes and compare results with analysis of other strains. Nevertheless, other strains with capsid sequences that closely match those of Mc10 and C12 are in the public database, which suggests the circulation of other recombinant sapovirus strains.
Recombination and evolution are important survival events for all living creatures as well as viruses. These events in viruses are not completely understood, but they can be potentially dangerous for host species, and they likely influence vaccine designs (15). From our studies, the human sapovirus and norovirus recombination appears limited to the intergenogroup because no intragenogroup or intragenus recombination has yet been identified that only occurs at the polymerase-capsid junction. Finally, the results of this study have increased our awareness of recombination in the Sapovirus genus and may have an influence on the future phylogenetic classification of sapovirus strains.
Dr. Katayama is a senior researcher at National Institute of Infectious Diseases, Tokyo. His research focuses on molecular epidemiologic studies on gastroenteritis viruses and sapovirus and norovirus genome expression.
Acknowledgment
This work was supported by grants-in-aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan and a Grant for Research on Re-emerging Infectious Diseases from the Ministry of Health, Labor, and Welfare of Japan. G.H. was supported by a Ph.D. scholarship from the Monbusho of Japan.
References
- Atmar RL, Estes MK. Diagnosis of noncultivatable gastroenteritis viruses, the human caliciviruses. Clin Microbiol Rev. 2001;14:15–37. DOIPubMedGoogle Scholar
- Noel JS, Fankhauser RL, Ando T, Monroe SS, Glass RI. Identification of a distinct common strain of “Norwalk-like viruses” having a global distribution. J Infect Dis. 1999;179:1334–44. DOIPubMedGoogle Scholar
- Chiba S, Nakata S, Numata-Kinoshita K, Honma S. Sapporo virus: history and recent findings. J Infect Dis. 2000;181(Suppl 2):S303–8. DOIPubMedGoogle Scholar
- Jiang X, Espul C, Zhong WM, Cuello H, Matson DO. Characterization of a novel human calicivirus that may be a naturally occurring recombinant. Arch Virol. 1999;144:2377–87. DOIPubMedGoogle Scholar
- Hansman GS, Katayama K, Maneekarn N, Peerakome S, Khamrin P, Tonusin S, Genetic diversity of norovirus and sapovirus in hospitalized infants with sporadic cases of acute gastroenteritis in Chiang Mai, Thailand. J Clin Microbiol. 2004;42:1305–7. DOIPubMedGoogle Scholar
- Katayama K, Shirato-Horikoshi H, Kojima S, Kageyama T, Oka T, Hoshino F, Phylogenetic analysis of the complete genome of 18 Norwalk-like viruses. Virology. 2002;299:225–39. DOIPubMedGoogle Scholar
- Lochridge VP, Hardy ME. Snow Mountain virus genome sequence and virus-like particle assembly. Virus Genes. 2003;26:71–82. DOIPubMedGoogle Scholar
- Vinje J, Green J, Lewis DC, Gallimore CI, Brown DW, Koopmans MP. Genetic polymorphism across regions of the three open reading frames of “Norwalk-like viruses.”. Arch Virol. 2000;145:223–41. DOIPubMedGoogle Scholar
- Worobey M, Holmes EC. Evolutionary aspects of recombination in RNA viruses. J Gen Virol. 1999;80:2535–43.PubMedGoogle Scholar
- Okada M, Shinozaki K, Ogawa T, Kaiho I. Molecular epidemiology and phylogenetic analysis of Sapporo-like viruses. Arch Virol. 2002;147:1445–51. DOIPubMedGoogle Scholar
- Lole KS, Bollinger RC, Paranjape RS, Gadkari D, Kulkarni SS, Novak NG, Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J Virol. 1999;73:152–60.PubMedGoogle Scholar
- Chen R, Neill JD, Noel JS, Hutson AM, Glass RI, Estes MK, Inter- and intragenus structural variations in caliciviruses and their functional implications. J Virol. 2004;78:6469–79. DOIPubMedGoogle Scholar
- Hansman G, Doan LP, Kguyen T, Okitsu S, Katayama K, Ogawa S, Detection of norovirus and sapovirus infection among children with gastroenteritis in Ho Chi Minh City, Vietnam. Arch Virol [serial on the Internet]. Jul 2004 [cited 2004 May 10]. Available from: http://springerlink.metapress.com/app/home/contribution.asp?wasp=n97bk9fqwgdrup9egxuq&referrer=parent&backto=issue,31,60;journal,1,92;linkingpublicationresults,1:100423,1
- Nilsson M, Hedlund KO, Thorhagen M, Larson G, Johansen K, Ekspong A, Evolution of human calicivirus RNA in vivo: accumulation of mutations in the protruding P2 domain of the capsid leads to structural changes and possibly a new phenotype. J Virol. 2003;77:13117–24. DOIPubMedGoogle Scholar
- Makeyev EV, Bamford DH. Evolutionary potential of an RNA virus. J Virol. 2004;78:2114–20. DOIPubMedGoogle Scholar
Figures
Cite This ArticleTable of Contents – Volume 10, Number 10—October 2004
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Grant S. Hansman, Department of Virology II, National Institute of Infectious Diseases. 4-7-1 Gakuen, Musashi-murayama, Tokyo 208-0011, Japan; fax: +81-42-565-3315
Top