Volume 19, Number 12—December 2013
Research
Zoonotic Chlamydiaceae Species Associated with Trachoma, Nepal
Abstract
Trachoma is the leading cause of preventable blindness. Commercial assays do not discriminate among all Chlamydiaceae species that might be involved in trachoma. We investigated whether a commercial Micro-ArrayTube could discriminate Chlamydiaceae species in DNA extracted directly from conjunctival samples from 101 trachoma patients in Nepal. To evaluate organism viability, we extracted RNA, reverse transcribed it, and subjected it to quantitative real-time PCR. We found that 71 (70.3%) villagers were infected. ArrayTube sensitivity was 91.7% and specificity was 100% compared with that of real-time PCR. Concordance between genotypes detected by microarray and ompA genotyping was 100%. Species distribution included 54 (76%) single infections with Chlamydia trachomatis, C. psittaci, C. suis, or C. pecorum, and 17 (24%) mixed infections that includied C. pneumoniae. Ocular infections were caused by 5 Chlamydiaceae species. Additional studies of trachoma pathogenesis involving Chlamydiaceae species other than C. trachomatis and their zoonotic origins are needed.
Trachoma was first recognized as an ocular disease in the 27th century BC in China (1). Subsequent reports documented the disease among the Egyptians and Greeks in the 19th and 1st centuries BC, respectively. The word trachoma derives from the Greek word for rough swelling, referring to the follicles that appear on the tarsal conjunctiva. Epidemic trachoma was spread from the Middle East to Europe during the Crusades and was a major cause of blindness during the Napoleonic era (1). The disease was eliminated from most industrialized countries after the industrial revolution, which heralded the institution of improved sanitation, hygiene, and nutrition. Currently, trachoma prevalence is hypoendemic, mesoendemic, and hyperendemic among populations residing in tropical developing countries.
During the past few decades, rates of trachoma have increased; in response, at the end of the 1990s, the World Health Organization developed the SAFE program with the goal of eliminating blinding trachoma by the year 2020. SAFE refers to Surgery, Antibiotics, Facial cleanliness, and Environmental improvements, specifically, surgery to correct trichiasis (in-turned eyelashes), oral antimicrobial drugs to treat Chlamydia trachomatis infections, facial cleanliness to decrease ocular infections, and environmental improvements such as latrines and wells to provide clean water. Unfortunately, most efforts have focused on the surgery and antimicrobial drug components and had disappointing results. Trichiasis often recurs months to years after surgery for 25%–75% of patients (2,3) and can be a result of reinfection (3). Infection often returns to pretreatment levels 6–24 months after termination of treatment (4,5). The recurrence of infection and disease is probably multifactorial. There is evidence that oral treatment of C. trachomatis infection blunts the immune response, increasing the patient’s susceptibility to reinfection (4). Furthermore, additional species of Chlamydiaceae, namely Chlamydia pneumoniae and C. psittaci, have been implicated in trachomatous disease by our group (6) and by another independent research group from Paris working in Guinea, Africa (7). To eliminate infections with species other than C. trachomatis, longer treatment intervals might be required (8).
Although some Chlamydiaceae screening tests and strain-typing methods exist, they are expensive, are time-consuming, require trained personnel, and are available only in specialized laboratories; most do not discriminate among species of Chlamydiaceae. The tests or methods include serotyping of the major outer membrane protein by using monoclonal or polyclonal antibodies that are species or genus specific; commercial nucleic acid amplification tests for C. trachomatis only (9); conventional species-specific and genus-specific PCRs (10); direct sequence analysis of ompA, 16S rRNS, or 23S rRNA genes (11,12); multilocus sequence typing for C. trachomatis (13, 14) and other species (14); real-time (RT)-PCR (6,15), multilocus variable number tandem repeat analysis (16); and the commercial micro ArrayTube or ArrayStrip (Alere Technologies, Jena, Germany) (17). Serotyping requires a cultured isolate, and techniques that involve sequencing might not be able to detect mixed-strain or mixed-species infections unless multiple strain–specific or species-specific primers are used, which require sufficient quantities of DNA. The advantage of the ArrayTube or ArrayStrip is that minimal DNA is required for amplification, and the hybridization patterns indicate species-specific nucleotide polymorphisms in regions of high sequence similarity.
The commercial ArrayTube assay has been successfully used to identify mixed infections among animals infected with multiple species of Chlamydiaceae (18,19). Because of these benefits, we investigated whether the ArrayTube could discriminate among Chlamydiaceae species in DNA that was extracted directly from conjunctival samples from trachoma patients residing in a trachoma-endemic region of Nepal. We also evaluated the correlation of the ArrayTube test with ompA genotypes. As an independent test for viability of Chlamydiaceae organisms, RNA was isolated from the same samples and tested by quantitative RT-PCR (qRT-PCR).
Study Population and Samples
We used a table of random numbers to randomly select 101 villagers, 1–65 years of age, who had follicular trachomatous inflammation and/or intense trachomatous inflammation and who resided in a trachoma-endemic region of the Lumbini Zone of southwestern Nepal. Patients were enrolled after they provided informed consent. For trachoma grading, we used the modified World Health Organization scale. Upper tarsal conjunctival samples were obtained by using dacron swabs (Hycor Biomedical, Portland, ME, USA), which were immediately placed in M4 transport media (Remel, Lenexa, KS, USA) and stored in liquid nitrogen as described (6). To avoid contamination, study personnel changed gloves between participants.
Ethics Statement
The study was approved by institutional review boards of the Nepal Netra Jhoti Shang (Kathmandu, Nepal) and the Children’s Hospital Oakland Research Institute (Oakland, CA, USA). Informed consent was obtained from each study participant. Oral consent was approved by both institutional review boards because of the high rate of illiteracy among the population. Consent was documented on the form by the team member who obtained the consent; the team member obtaining the consent signed the form stating that consent had been obtained. Since some study participants were minors, parents provided consent for their child to participate.
RNA and DNA Purification
Genomic DNA was extracted from the conjunctival swab samples by using the Roche High Pure kit (Roche, Pleasanton, CA, USA), and RNA was extracted by using the RNeasy kit (QIAGEN, Valencia, CA, USA) as described (6,20). RNA was reverse transcribed to cDNA by using a TaqMan reverse transcription kit (Applied Biosystems, Foster City, CA, USA) as described (6,20). DNA and RNA were stored at −80°C until use.
DNA Microarray Assay
To examine samples for the presence of any of the 9 Chlamydiaceae species and Waddlia chondrophila and Simkania negevensis, we performed the ArrayTube assay as described (17,19). Briefly, DNA from each sample was amplified and biotin labeled in 40 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 30 s by using primers U23F-19 and 23R-22 (Table 1). Hybridization was conducted in the ArrayTube vessel at 58°C for 1 h. After 3 wash steps, hybridization signals were visualized by using streptavidin-conjugated peroxidase-catalyzed precipitation. The resulting patterns were processed by using the ATR-01 ArrayTube reader (Alere Technologies) and the Iconoclust 2.3 program (Alere).
Genus-Specific and Species-Specific qRT-PCR and ompA Genotyping
qRT-PCR was conducted by using genus-specific and species-specific primers (Table 1) along with appropriate controls including β-actin as described (6,20). Briefly, each reaction contained 1X SYBR Green PCR MasterMix (Applied Biosystems), 300 nmol/L of each primer, and 5 μL of sample DNA in a volume of 25 μL in duplicate in a 96-well plate. The thermocycling consisted of 10 min at 94°C followed by 40 cycles of 15 s at 94°C and 1 min at 60°C. Samples that were positive by qRT-PCR were subjected to PCR with 16S rRNA genus-specific primers in addition to species-specific ompA primers (Table 1). The PCR reagents, controls, thermocycling, and sequencing by BigDye Terminator (Applied Biosystems) automated capillary sequencing were used or performed as described (6,20). In addition, ompA genotyping of samples showing monoinfection with C. trachomatis was conducted by using the ArrayStrip microarray assay as described (21).
Data Analysis
Outcome variables included single or mixed infection, Chlamydiaceae species causing the infection, and ompA genotype. The association between discrete variables was analyzed by using the Fisher exact test or the Pearson χ2 test by Stata 10 (College Station, TX, USA). A p value of <0.05 was used as the cutoff for determining statistical significance.
The distribution of villagers by age and sex is shown in Table 2; none lived in the same household. The rate of mixed infections was significantly higher for female than for male participants (p = 0.0011), although the overall rates of infection did not differ. Single or mixed infections did not differ by age group, although the rate of single and mixed infections among those >10 years of age was significantly higher (p = 0.0472). The mean age was 26 years for female and 28 years for male participants.
For purposes of cross-comparison, the ArrayTube and qRT-PCR analyses were run independently. For the 101 samples, the ArrayTube had a sensitivity of 91.7% and a specificity of 100% compared with sensitivity and specificity of qRT-PCR (Technical Appendix Table). None of the samples yielded positive results for C. abortus by ArrayTube or qRT-PCR. Six samples were not detected by the ArrayTube but were C. trachomatis positive by qRT-PCR. All samples positive by qRT-PCR were genotyped; the ompA genotypes matched 100% of those identified by ArrayTube.
Figure 1
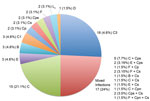
Figure 1. . . Chlamydiaceae infections among 101 villagers residing in a trachoma-endemic region of southwestern Nepal identified by the ArrayTube (Alere Technologies, Jena, Germany), real-time PCR, and ompA genotyping. The number and...
Figure 2

Figure 2. . . Identification of Chlamydiaceae triple infection by using the ArrayTube (Alere Technologies, Jena, Germany) assay. A) Biotinylated PCR product from a DNA extract was hybridized to a DNA microarray carrying...
Of the 101 participating villagers, 71 (70.3%) were infected (Figure 1); 26 (37%) of the infections involved a single or mixed infection with a species other than C. trachomatis or in combination with C. trachomatis. The 54 (76%) single infections were 48 (88.9%) C. trachomatis, 2 (3.7%) C. psittaci, 2 (3.7%) C. suis, and 2 (3.7%) C. pecorum. Most infections were caused by C. trachomatis strains C, C1, and C3, although urogenital strains B, D, E, F, and L2 were also represented. The 17 (24%) mixed infections were 15 (88.2%) C. trachomatis plus another species and 2 (11.8%) other species. Specifically, they were 7 (41.2%) C. trachomatis plus C. psittaci, 5 (29.4%) C. trachomatis plus C. suis, 2 (11.7%) C. psittaci plus C. suis, 1 (5.9%) C. trachomatis plus C. pneumoniae, 1 (5.9%) C. trachomatis plus C. pecorum, and 1 (5.9%) C. psittaci plus C. suis and C. trachomatis. There were no statistically significant differences by patient age or sex for infections caused by C. trachomatis or other species. Figure 2, panel A, shows the ArrayTube assay for the sample from the patient infected with the latter 3 species. Figure 2, panel B, shows an example of microarray-based ompA genotyping.
Although C. trachomatis is well established as a cause of trachoma (1), the high percentage of infections with other species (37%) found in this study suggests a role for these pathogens in trachoma pathogenesis. In addition, the use of 2 independent methods to detect Chlamydiaceae, one of which used RNA to demonstrate the presence of viable Chlamydiaceae species in trachomatous eyes, lends further support to this theory. Our findings also support 2 previous studies; 1 conducted by our group and 1 conducted by an independent research group in Paris. Our group detected C. trachomatis, C. psittaci, and C. pneumoniae DNA and RNA among Nepali villagers in all age groups (6). Just under 60% of the cases were caused by single or mixed infections with species other than C. trachomatis. In addition, infection with each species was significantly associated with antibodies to chlamydial heat shock protein 60, a known virulence factor for the organism associated with inflammation and trachomatous scarring. The Paris group conducted their study in a trachoma-endemic region of Guinea, Africa, and detected C. trachomatis, C. psittaci, and C. pneumoniae among children 1–10 years of age by using RT-PCR that targeted a conserved sequence of 16S rRNA (7).
These findings are not surprising for several reasons. Historically, swab samples from persons in a trachoma-endemic area were cultured and then serologically typed by using microimmunofluorescence, a technique that requires technical expertise, and only 1 strain each of C. pneumoniae and C. psittaci was included. However, cross-reactivity among species has been observed (22). Immunoreactivity to other species, therefore, might not be considered as evidence for infection with these pathogens. However, in a study in Sudan where trachoma is hyperendemic, immunoreactivity to all 3 species was found among 3% of persons with clinical trachoma (23). Their findings suggest either cross-reactivity or infection with these species. Currently, conjunctival swab samples from patients in trachoma-endemic areas are usually only tested by C. trachomatis–specific commercial nucleic acid amplification tests or by C. trachomatis–specific in-house PCR. Although a pan-Chlamydiales PCR is available, it might not detect mixed infections because an additional cloning step is required (24). Furthermore, this test has not been applied to samples from persons in a trachoma-endemic area. Consequently, unless investigators think to use specific tests to look for other species, they will not be detected.
Our study identified ocular infections with 5 Chlamydiaceae species among trachoma patients. Over the past 5 years, improved technology has enabled increasing identification of single and mixed infections with Chlamydiaceae species among mammals and birds (10,19,25). Co-infections with C. psittaci and C. abortus have been found in cow milk, and co-infections with C. abortus and C. pecorum have been identified in conjunctival and nasal swab specimens from calves (19). Mixed infections among bovine abortion cases have included C. abortus and C. suis (25). When cloacal swab samples and fecal samples from pigeons were tested, mixed infections with C. psittaci, combined with C. pecorum, C. abortus, or C. trachomatis, along with unclassified Chlamydiaceae spp., were discovered (18). Most studies used the commercial ArrayTube or ArrayStrip.
Prevalence of ocular infections with Chlamydiaceae species in different mammals is high, which supports the notion that humans also are probably susceptible to ocular infection and disease with zoonotic species. A recent study identified several Chlamydiaceae species infecting the diseased and healthy eyes of sheep; these species were C. abortus, C. pecorum, C. suis, and uncultured Chlamydia-like organisms (26). C. psittaci has been associated with ocular diseases in sheep and koalas (27). C. suis is also well known as a cause of conjunctivitis among pigs (28), and C. pecorum is associated with keratoconjunctivitis among sheep and goats (26). One study tracked Chlamydiaceae infections among humans and their domesticated animals and found C. psittaci on ocular swabs from humans, cattle, buffaloes, sheep, and goats inhabiting the same compounds (29).
The plethora of mixed infections, along with single infections with various Chlamydiaceae species, among domesticated animals (such as ducks, pigs, cows, sheep, goats, horses, and cattle) suggests multiple opportunities for transmission to humans and development of disease. C. pneumoniae (30) and C. psittaci (31,32) are known to be transmissible from human to human. Alternatively, infection and reinfection from animals could be another mechanism for transmission in which human-to-human transmission, which is necessary for the human-confined pathogen C. trachomatis, would not be required. In addition, clinical outcomes could be worse when caused by mixed chlamydial infections rather than monoinfection, as was suggested in a study of ovine abortion (33).
A major question is whether C. trachomatis strains that cause sexually transmitted diseases (STDs) or Chlamydiaceae species that cause zoonotic infection are capable of causing chronic infection and the cycles of reinfection that are characteristic of trachoma caused by C. trachomatis. STD-strain infections among children causing trachoma-like disease have been reported. In a study by Harrison et al. (34), urogenital strain J was isolated from the conjunctiva of a Navajo child who had trachoma. This strain was also noted to have been isolated from the urogenital tract of Native American women in the same area. Mordhorst et al. (35) isolated strains Ba, D, E, G, and H from 14 patients in Denmark for whom onset of infection and trachoma occurred during childhood. Of these 14 patients, trachomatous disease was severe for 5. We have also documented trachoma-like disease caused by C. psittaci and C. pneumoniae in the United States (8). Another report notes the isolation of C. pneumoniae from a laboratory technician with acute follicular conjunctivitis who had been working with the agent (36). Incidents of ocular infection with C. suis have occurred among pig farm and slaughterhouse workers (D. Vanrompay, pers. comm.); however, these infections were asymptomatic.
Animal models of trachoma also lend support to the pathogenic role of STD strains and other zoonotic Chlamydiaceae species in trachoma. STD strains E and G have been shown to produce severe ocular disease similar to trachoma in macaque or baboon models of trachoma (37–39). There is also a guinea pig model of trachoma in which C. caviae has been shown to produce pathologic changes similar to those of trachoma (40). These collective findings indicate that multiple zoonotic species can probably infect the eyes of humans and might contribute to trachomatous disease pathogenesis.
In our study, we randomly selected villagers with follicular or intense trachomatous inflammation to screen for Chlamydiaceae infections. None of the selected villagers lived in the same household. Most infections were with trachoma C strains, although urogenital strains B, D, E, F, and L2 were also detected. We have documented urogenital infections in trachoma patients in Nepal but not with L2 (6), perhaps because of the small sample size. We identified 4 other zoonotic species as etiologic agents in single or mixed infections. These species are common among domesticated animals (such as pigeons, pigs, cows, and buffaloes), which in trachoma-endemic communities are commonly kept for consumption or agricultural purposes. In our previous study (6), we tested for only C. psittaci and C. pneumoniae in addition to C. trachomatis.
The mixed infection rate of 24% found in our study was comparable to the rate of 35% found in our previous study, although the higher rate might have reflected the testing of multiple family members who would have had the same exposure to infected animals (6). The significantly higher rate of mixed infections among female participants is consistent with their societal role of caring for domesticated animals and the enhanced opportunity for contact with potentially infected animals. No significant associations were found between age or sex and infecting species. This finding is similar to that of the previous study (6), indicating that these species are probably prevalent in the communities and that all villagers are susceptible to zoonotic infection.
The ArrayTube was 91.7% sensitive and 100% specific compared with qRT-PCR. That 5 samples were negative by the ArrayTube might reflect the higher sensitivity of amplifying a single target in qRT-PCR compared with amplifying multiple targets in a single ArrayTube assay. Nonetheless, the ArrayTube assay is a relatively quick assay for screening populations in trachoma-endemic areas for Chlamydiaceae species. It is ideally suited for detecting mixed infections that might be missed by tests that target a single species, that amplify only the most abundant species in the sample, or that require additional DNA for multiple strain-specific or species-specific amplifications.
Identifying Chlamydiaceae species distribution among persons in a trachoma-endemic area is critical for understanding disease prevalence and instituting appropriate therapeutic regimens for the specific species (8). For assessment of the prevalence of infections caused by all Chlamydiaceae species and for a better understanding of their zoonotic origins, additional studies using the ArrayTube are warranted in other trachoma-endemic countries worldwide. For prevention of transmission from animal to human hosts, interventions will probably need to be instituted. Our results, then, represent findings that could help guide the World Health Organization program for eliminating blinding trachoma by the year 2020. Finally, understanding the full effects of multiple Chlamydiaceae species on the epidemiology, immunopathology, and disease outcome of trachoma will be a major research challenge. Although additional studies are needed, on the basis of our findings, vaccine design will probably need to consider the potential diversity of the host immune response to different Chlamydiaceae pathogens.
Dr Dean is director of the Children’s Global Health Initiative and a senior scientist in the Center for Immunobiology and Vaccine Development at the Children’s Hospital Oakland Research Institute, Oakland. She is also a faculty member in the graduate program in bioengineering at the University of California, San Francisco, and the University of California, Berkeley. Her research interests focus on the pathogenesis and comparative genomics and molecular epidemiology of chlamydial ocular and sexually transmitted diseases.
Acknowledgments
We thank the villagers of Kapilvastu District, Nepal, for their invaluable participation in the project.
This research was funded in part by the National Science Foundation–US Department of Agriculture grant 2009-65109-05760 (to D.D.) and Public Health Service grant R01 AI098843 from the National Institutes of Health.
References
- Dean D. Pathogenesis of chlamydial ocular infections. In: Tasman W, Jaeger EA, editors. Duane's foundations of clinical ophthalmology. Philadelphia: Lippincott Williams & Wilkins; 2010. p. 678–702.
- Zhang H, Kandel RP, Sharma B, Dean D. Risk factors for recurrence of postoperative trichiasis: implications for trachoma blindness prevention. Arch Ophthalmol. 2004;122:511–6. DOIPubMedGoogle Scholar
- Zhang H, Kandel RP, Atakari HK, Dean D. Impact of oral azithromycin on recurrence of trachomatous trichiasis in Nepal over 1 year. Br J Ophthalmol. 2006;90:943–8. DOIPubMedGoogle Scholar
- Atik B, Thanh TT, Luong VQ, Lagree S, Dean D. Impact of annual targeted treatment on infectious trachoma and susceptibility to reinfection. JAMA. 2006;296:1488–97. DOIPubMedGoogle Scholar
- Chidambaram JD, Alemayehu W, Melese M, Lakew T, Yi E, House J, Effect of a single mass antibiotic distribution on the prevalence of infectious trachoma. JAMA. 2006;295:1142–6. DOIPubMedGoogle Scholar
- Dean D, Kandel RP, Adhikari HK, Hessel T. Multiple Chlamydiaceae species in trachoma: implications for disease pathogenesis and control. PLoS Med. 2008;5:e14. DOIPubMedGoogle Scholar
- Goldschmidt P, Rostane H, Sow M, Goepogui A, Batellier L, Chaumeil C. Detection by broad-range real-time PCR assay of Chlamydia species infecting human and animals. Br J Ophthalmol. 2006;90:1425–9. DOIPubMedGoogle Scholar
- Lietman T, Brooks D, Moncada J, Schachter J, Dawson C, Dean D. Chronic follicular conjunctivitis associated with Chlamydia psittaci or Chlamydia pneumoniae. Clin Infect Dis. 1998;26:1335–40. DOIPubMedGoogle Scholar
- Gaydos CA. Nucleic acid amplification tests for gonorrhea and Chlamydia: practice and applications. Infect Dis Clin North Am. 2005;19:367–86. DOIPubMedGoogle Scholar
- Sachse K, Vretou E, Livingstone M, Borel N, Pospischil A, Longbottom D. Recent developments in the laboratory diagnosis of chlamydial infections. Vet Microbiol. 2009;135:2–21. DOIPubMedGoogle Scholar
- Dean D, Patton M, Stephens RS. Direct sequence evaluation of the major outer membrane protein gene variant regions of Chlamydia trachomatis subtypes D′, I′, and L2′. Infect Immun. 1991;59:1579–82 .PubMedGoogle Scholar
- Geens T, Desplanques A, Van Loock M, Bonner BM, Kaleta EF, Magnino S, Sequencing of the Chlamydophila psittaci ompA gene reveals a new genotype, E/B, and the need for a rapid discriminatory genotyping method. J Clin Microbiol. 2005;43:2456–61. DOIPubMedGoogle Scholar
- Dean D, Bruno WJ, Wan R, Gomes JP, Devignot S, Mehari T, Predicting phenotype and emerging strains among Chlamydia trachomatis infections. Emerg Infect Dis. 2009;15:1385–94. DOIPubMedGoogle Scholar
- Pannekoek Y, Dickx V, Beeckman DS, Jolley KA, Keijzers WC, Vretou E, Multi locus sequence typing of Chlamydia reveals an association between Chlamydia psittaci genotypes and host species. PLoS ONE. 2010;5:e14179. DOIPubMedGoogle Scholar
- Pantchev A, Sting R, Bauerfeind R, Tyczka J, Sachse K. Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp Immunol Microbiol Infect Dis. 2010;33:473–84. DOIPubMedGoogle Scholar
- Bom RJ, Christerson L, Schim van der Loeff MF, Coutinho RA, Herrmann B, Bruisten SM. Evaluation of high-resolution typing methods for Chlamydia trachomatis in samples from heterosexual couples. J Clin Microbiol. 2011;49:2844–53. DOIPubMedGoogle Scholar
- Sachse K, Hotzel H, Slickers P, Ellinger T, Ehricht R. DNA microarray-based detection and identification of Chlamydia and Chlamydophila spp. Mol Cell Probes. 2005;19:41–50 and. DOIPubMedGoogle Scholar
- Sachse K, Kuehlewind S, Ruettger A, Schubert E, Rohde G. More than classical Chlamydia psittaci in urban pigeons. Vet Microbiol. 2012;157:476–80. DOIPubMedGoogle Scholar
- Borel N, Kempf E, Hotzel H, Schubert E, Torgerson P, Slickers P, Direct identification of chlamydiae from clinical samples using a DNA microarray assay: a validation study. Mol Cell Probes. 2008;22:55–64. DOIPubMedGoogle Scholar
- Gomes JP, Hsia RC, Mead S, Borrego MJ, Dean D. Immunoreactivity and differential developmental expression of known and putative Chlamydia trachomatis membrane proteins for biologically variant serovars representing distinct disease groups. Microbes Infect. 2005;7:410–20. DOIPubMedGoogle Scholar
- Ruettger A, Feige J, Slickers P, Schubert E, Morre SA, Pannekoek Y, Genotyping of Chlamydia trachomatis strains from culture and clinical samples using an ompA-based DNA microarray assay. Mol Cell Probes. 2011;25:19–27. DOIPubMedGoogle Scholar
- Kern DG, Neill MA, Schachter J. A seroepidemiologic study of Chlamydia pneumoniae in Rhode Island. Evidence of serologic cross-reactivity. Chest. 1993;104:208–13. DOIPubMedGoogle Scholar
- Mahmoud E, Elshibly S, Mardh PA. Seroepidemiologic study of Chlamydia pneumoniae and other chlamydial species in a hyperendemic area for trachoma in the Sudan. Am J Trop Med Hyg. 1994;51:489–94 .PubMedGoogle Scholar
- Lienard J, Croxatto A, Aeby S, Jaton K, Posfay-Barbe K, Gervaix A, Development of a new Chlamydiales-specific real-time PCR and its application to respiratory clinical samples. J Clin Microbiol. 2011;49:2637–42 and. DOIPubMedGoogle Scholar
- Blumer S, Greub G, Waldvogel A, Hassig M, Thoma R, Tschuor A, Waddlia, Parachlamydia and Chlamydiaceae in bovine abortion. Vet Microbiol. 2011;152:385–93. DOIPubMedGoogle Scholar
- Polkinghorne A, Borel N, Becker A, Lu ZH, Zimmermann DR, Brugnera E, Molecular evidence for chlamydial infections in the eyes of sheep. Vet Microbiol. 2009;135:142–6. DOIPubMedGoogle Scholar
- Girjes AA, Hugall A, Graham DM, McCaul TF, Lavin MF. Comparison of type I and type II Chlamydia psittaci strains infecting koalas (Phascolarctos cinereus). Vet Microbiol. 1993;37:65–83. DOIPubMedGoogle Scholar
- Becker A, Lutz-Wohlgroth L, Brugnera E, Lu ZH, Zimmermann DR, Grimm F, Intensively kept pigs pre-disposed to chlamydial associated conjunctivitis. J Vet Med A Physiol Pathol Clin Med. 2007;54:307–13. DOIPubMedGoogle Scholar
- Osman KM, Ali HA, Eljakee JA, Galal HM. Prevalence of Chlamydophila psittaci infections in the eyes of cattle, buffaloes, sheep and goats in contact with a human population. Transbound Emerg Dis. 2013;60:245–51 .DOIPubMedGoogle Scholar
- Yamazaki T, Nakada H, Sakurai N, Kuo CC, Wang SP, Grayston JT. Transmission of Chlamydia pneumoniae in young children in a Japanese family. J Infect Dis. 1990;162:1390–2. DOIPubMedGoogle Scholar
- Hughes C, Maharg P, Rosario P, Herrell M, Bratt D, Salgado J, Possible nosocomial transmission of psittacosis. Infect Control Hosp Epidemiol. 1997;18:165–8. DOIPubMedGoogle Scholar
- Lenzko H, Moog U, Henning K, Lederbach R, Diller R, Menge C, High frequency of chlamydial co-infections in clinically healthy sheep flocks. BMC Vet Res. 2011;7:29. DOIPubMedGoogle Scholar
- Harrison HR, Boyce WT, Wang SP, Gibb GN, Cox JE, Alexander ER. Infection with Chlamydia trachomatis immunotype J associated with trachoma in children in an area previously endemic for trachoma. J Infect Dis. 1985;151:1034–6. DOIPubMedGoogle Scholar
- Mordhorst CH, Wang SP, Grayston JT. Childhood trachoma in a nonendemic area. Danish trachoma patients and their close contacts, 1963 to 1973. JAMA. 1978;239:1765–71. DOIPubMedGoogle Scholar
- Forsey T, Darougar S. Acute conjunctivitis caused by an atypical chlamydial strain: Chlamydia IOL 207. Br J Ophthalmol. 1984;68:409–11. DOIPubMedGoogle Scholar
- Wang S, Grayston JT. Trachoma in the Taiwan monkey Macaca cyclopis. Ann N Y Acad Sci. 1962;98:177–87. DOIPubMedGoogle Scholar
- Taylor HR, Prendergast RA, Dawson CR, Schachter J, Silverstein AM. An animal model for cicatrizing trachoma. Invest Ophthalmol Vis Sci. 1981;21:422–33 .PubMedGoogle Scholar
- Collier LH. Experimental infection of baboons with inclusion blennorrhea and trachoma. Ann N Y Acad Sci. 1962;98:188–96. DOIPubMedGoogle Scholar
- Watkins NG, Hadlow WJ, Moos AB, Caldwell HD. Ocular delayed hypersensitivity: a pathogenetic mechanism of chlamydial-conjunctivitis in guinea pigs. Proc Natl Acad Sci U S A. 1986;83:7480–4. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 19, Number 12—December 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Deborah Dean, Children’s Hospital Oakland Research Institute, 5700 Martin Luther King Jr. Way, Oakland, CA 94609, USADeborah Dean, Children’s Hospital Oakland Research Institute, 5700 Martin Luther King Jr. Way, Oakland, CA 94609, USA
Top