Volume 6, Number 6—December 2000
Research
Molecular Evidence of Clonal Vibrio parahaemolyticus Pandemic Strains
Abstract
The upsurge in worldwide incidence of Vibrio parahaemolyticus infection in the last 5 years has been attributed to the recent appearance of three serotypes with pandemic potential: O3:K6, O4:K68, and O1:K untypeable (KUT). Thirty-five strains of these serotypes, isolated from different countries over 4 years, were characterized by ribotyping and pulsed-field gel electrophoresis to determine their origin. The ribotypes of the strains of these serotypes were indistinguishable, except for a Japanese tdh- negative O3:K6 strain and a U.S. clinical O3:K6 isolate, which had slightly different profiles. The migration patterns of the NotI-digest of the total DNA of the strains were similar, and only slight variations were observed between the serotypes. By contrast, the O3:K6 and O1:KUT strains isolated before 1995 and strains of other serotypes had markedly different profiles. The O4:K68 and O1:KUT strains most likely originated from the pandemic O3:K6 clone.
Infections caused by Vibrio parahaemolyticus, a halophilic member of the genus Vibrio, have increased globally in the last 5 years. V. parahaemolyticus diarrhea results from consuming raw or undercooked seafoods, although other routes of transmission have been documented (1). Studies implicate the thermostable direct hemolysin (TDH) and the TDH-related hemolysin (TRH), encoded by the tdh and trh gene, respectively, as the major virulence factors of this organism (2-5). Therefore, the presence of the tdh gene marked by a β-type hemolysis on Wagatsuma agar (2,3), the trh gene correlated to a positive urease test (6), or both serve as markers for pathogenic strains. Recently, three major serotypes--O3:K6, O4:K68, and O1:K untypeable (KUT), listed in chronological order of appearance-- have caused a pandemic of V. parahaemolyticus infection (7-10). Strains of these serotypes have been responsible for gastroenteritis in India, other Southeast Asian countries, and the United States (9-12). In Calcutta, strains of the O3:K6 serotype have been responsible for the high incidence of V. parahaemolyticus-mediated gastroenteritis since February 1996 (7). Likewise, this serotype was isolated in other Asian countries (including Laos, Taiwan, and Japan) and the United States.
This alarming rise of a serotype previously associated with only sporadic cases of gastroenteritis was monitored closely. The O3:K6 strains isolated before 1995 (henceforth referred to as old O3:K6) and those isolated since 1995 (referred to as new O3:K6) were analyzed for variation in nucleotide sequence of the toxRS region and differed invariably in seven bases within the 1,364-bp toxRS region (10). Two out of the seven unique bases were exploited to develop the group-specific polymerase chain reaction (GS-PCR) that distinguished between the new and the old O3:K6 isolates. Examination of the non-O3:K6 isolates by GS-PCR showed that the toxRS sequences of the recent isolates of O4:K68 and O1:KUT serotypes were identical to those of the new O3:K6 isolates (10). All the strains of the new clone (regardless of serotype or place of isolation) carried the tdh gene but not the trh gene. These strains also exhibited a unique arbitrarily primed PCR profile distinct from the strains of other serotypes, supporting the hypothesis that the O4:K68 and O1:KUT strains evolved from the newly emerged O3:K6 clone. Using ribotyping and pulsed-field gel electrophoresis (PFGE), we examined whether a single clone expressed as three different serotypes of V. parahaemolyticus is causing the pandemic.
Bacterial Strains
A total of 35 pandemic strains of V. parahaemolyticus (21 of O3:K6, 10 of O4:K68, and 4 of O1:KUT) isolated from 1996 to 1999 from widely separated geographic regions were examined by ribotyping and PFGE (Table 1). Thirteen nonpandemic strains of O3:K6, O1:KUT, and other serotypes isolated before February 1996 were included as external controls (Table 2).
Purification of Genomic DNA
A modification of the method of Murray and Thompson (13) was used to extract chromosomal DNA. Briefly, cells from 18-hour culture in Luria-Bertani (LB) broth, Miller (Difco Laboratories, Detroit, MI) with 3% NaCl, were harvested by centrifugation at 6,000 x g for 5 minutes. The pelleted cells were resuspended in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0), treated with 10% sodium dodecyl sulfate (SDS) and proteinase K and incubated for 1 hours at 37°C. After incubation, CTAB/NaCl (10% cetyl trimethyl ammonium bromide in 0.7 M NaCl) was added and incubated at 65°C for 10 minutes. The aqueous phase was then treated with phenol-chloroform, and the DNA was pelleted and washed with 70% ethanol. Purified DNA was suspended in TE buffer and treated with RNase at 37°C for 30 minutes.
Ribotyping
Restriction enzyme BglI (Boehringer GmbH, Mannheim, Germany) was used for ribotyping of the V. parahaemolyticus strains. The genomic fragments were electrophoresed on a 1% SeaKem agarose gel (FMC Bioproducts, Rockland, ME) using Tris-acetate (TAE) buffer (0.04 M TAE, 0.001 M EDTA [pH 8.0]). For Southern blotting, the gel was treated successively in 0.25 N HCl for 10 minutes to allow partial depurination and cleavage of large fragments, in denaturation solution composed of 0.5 M NaOH, 1.5 M NaCl for 30 minutes and in 0.5 M Tris-HCl (pH 7.4) for 30 minutes. DNA was then transferred to Hybond N+ membrane (Amersham International PLC, Buckinghamshire, England), using 20X SSC (3 M sodium chloride, 0.3 M sodium citrate; by vacuum blotter (Pharmacia, Sweden). The membrane was then washed with 20X SSC and dried at room temperature followed by fixation in a UV cross-linker (Bio-Rad Laboratories, Richmond, CA). A 7.5-kb BamHI fragment of the recombinant plasmid pKK3535 containing an rRNA operon of Escherichia coli (14) was used as the rrn gene probe for ribotyping. Labeling of the probes, hybridization, and detection of bands were carried out according to the instructions of the manufacturer of the ECL detection system (Amersham Life Science, UK).
DNA Extraction and Digestion for PFGE
The test strains grown on LB agar, Miller (Difco) with 3% NaCl were transferred to 3 mL LB broth, Miller (Difco) with 3% NaCl and cultured overnight at 37°C with shaking at 100 rpm. One hundred microliters of the overnight culture was transferred to 8 mL of LB broth, Miller and incubated at 37°C with shaking at 100 rpm until the culture attained an optical density of 0.9 at 600 nm. Bacterial cells were harvested from 1 mL of the culture by centrifugation and resuspended in 0.5 mL cell lysis buffer (10 mM Tris-HCl [pH7.2], 20 mM NaCl, 50 mM EDTA). Agarose plugs were prepared by mixing equal volumes of bacterial suspension with 2% low-melting agarose. The bacterial cells in the agarose plugs were lysed by treatment with lysis solution (1 mg of lysozyme per mL, 0.4% N-sodium lauryl sarcosine, 0.2% Na-deoxycholate, 10 mM Tris [pH 7.2], 50 mM NaCl) at 37°C overnight and treated with proteinase K at 50°C overnight. The plugs were washed with washing buffer containing 20 mM Tris HCl (pH 8.0) and 50 mM EDTA. Agarose plugs containing genomic DNA were equilibrated in enzyme buffer for 1 hour at room temperature and were cleaved in 600 µL of enzyme buffer H (500 mM Tris-HCl [pH7.5], 100 mM MgCl2 , 10 mM DTT, 100 mM NaCl) containing 50 U of NotI enzyme at 37°C overnight.
PFGE
PFGE of NotI-digested inserts was performed on 1% agarose (Bio-Rad) by the contour-clamped homogenous electric field method on a CHEF Mapper system (Bio-Rad) in 0.5X TBE buffer (44.5 mM Tris-HCl, 44.5 mM boric acid, 1.0 mM EDTA, pH 8.0) for 40 hours, 24 minutes. A DNA size standard (Bacteriophage λ-ladder, Bio-Rad) was used as the molecular mass standard, and a minichiller (Bio-Rad) was used to maintain the temperature of the buffer at 14°C. Run conditions were generated by the autoalgorithm mode of the CHEF Mapper PFGE system by using a 20- to 300-kb size range. After electrophoresis, the gel was stained in ethidium bromide (1 µg/mL) for 30 minutes and destained in water for 15 minutes twice. The DNA bands were visualized and photographed with the Gel Doc 2000 (Bio-Rad) (8).
Figure 1
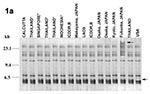
Figure 1. Southern blot hybridization of BglI-digested Vibrio parahaemolyticus chromosomal DNA with rRNA probe. 1a-1c, ribotype patterns of the O3:K6, O4:K68, and O1:K untypeable (KUT) strains, respectively, isolated from different countries. 1d, ribotype...
The organization of the rrn genes in the chromosomes of V. parahaemolyticus was examined by Southern hybridization of BglI-digested total DNA with rRNA-specific DNA probe. The ribotyping patterns observed with the strains of the three different serotypes are shown in Figure 1. The DNA probe specific to rrn genes hybridized with 11 fragments, 23.0 kb to 4.0 kb. Strains of the new O3:K6, O4:K68, and O1:KUT serotypes exhibited the R4 ribosomal banding pattern, which was the major pattern exhibited by 76% of the O3:K6 isolates in India from February to August 1996 (8). However, the O3:K6 strain, FIHES98VI-32-4, isolated in Japan, which was devoid of the tdh gene and was included in this study because it was positive by GS-PCR (10), differed in the 23.1-kb region. Another O3:K6 strain, BE98-2062, isolated from the United States, differed from the other strains by a single band near the 6.0-kb region. Representative nonpandemic strains--including the old O3:K6, O1:KUT, and other unrelated serotypes--were similarly analyzed for the organization of the rrn genes (Figure 1d). The nonpandemic strains showed heterogenous ribotype profiles, each pattern markedly different from the other and from that exhibited by the pandemic strains.
Figure 2

Figure 2. Electrophoretic migration pattern of the NotI-digested Vibrio parahaemolyticus genomic DNA obtained by pulsed-field gel electrophoresis (PFGE). 2a-2c, PFGE patterns of the O3:K6, O4:K68, and O1:K untypeable (KUT) strains, respectively, isolated from...
The ribotyping data were substantiated by studying the electrophoretic migration pattern of NotI-digested fragments of the chromosomal DNA of each of the 35 pandemic strains of V. parahaemolyticus obtained by PFGE. The PFGE data for the O3:K6, O4:K68, and O1:KUT strains are shown in Figure 2. Except for the tdh-negative O3:K6 strain from Japan (FIHES98VI-32-4), all the O3:K6 strains show almost identical RFLP patterns. The O3:K6 strain, BE98-2062, isolated in the United States, showed minor variation from the other strains near the 58.5-kb region. The PFGE patterns of all O4:K68 strains isolated from different geographic locations were identical and nearly similar to the pattern obtained with the O3:K6 strains (Figure 2b). Strains of this serotype, however, differ from the O3:K6 strains by one band near the 240-kb region and by the higher intensity of a band at the 194-kb region, which could indicate the comigration of more than one band. Similarly, although all four O1:KUT strains were identical to each other, they differed from the O4:K68 serotype by the absence of a single band in the 200-kb region and from the O3:K6 serotype by the absence of bands at the 240-kb and 200-kb regions (Figure 2c).
Since all isolates of the three different serotypes, except FIHES98VI-32-4, varied by one or two bands, all appear to have originated from a common ancestor. The variation may be due to the difference in the genetic organization of the O and K antigen biosynthesis gene clusters in the strains. The banding pattern of the NotI-digested DNA fragments, obtained by PFGE of the nonpandemic strains, is shown in Figure 2d. Considerable polymorphism was observed between the pandemic and nonpandemic strains and between the nonpandemic strains of various serotypes.
Until recently, V. parahaemolyticus caused sporadic and localized diarrhea and--unlike toxigenic V. cholerae O1 and O139--was never associated with a pandemic. However, with the advent of the new O3:K6 strains in 1996, the epidemiology of this organism abruptly changed. The dominant and continued occurrence of this serotype was reported from eight countries. The extent and rapidity of spread of the new O3:K6 strains signaled the beginning of the first pandemic of V. parahaemolyticus. We have shown that the recent O3:K6 isolates from eight countries were identical in the RFLP of the rRNA genes and showed similar PFGE profiles. We have also shown that strains of two other serotypes, O4:K68 and O1:KUT, isolated since 1995 possessed ribotype and PFGE patterns similar to those of the new O3:K6 strains. Variations between the three pandemic serotypes are minor when compared to the differences seen with the nonpandemic strains. Hence, from the molecular analysis and chronology of appearance of these strains, the O4:K68 and O1:KUT isolates appear to have originated from the existing O3:K6 clone. The ribotype and PFGE patterns displayed by the pandemic clone are unique. Therefore, a single clone may be responsible for the emergence of pandemic serotypes that have different somatic and capsular antigens. This study suggests that the epidemiologically related strains may also be genetically related. The abrupt origin of this pandemic clone and, more importantly, the sudden acquisition of pandemic properties by three different serotypes of V. parahaemolyticus, almost 5 decades after its discovery, require further scrutiny.
Ms. Chowdhury is a senior research fellow under the Indian Council of Medical Research. She is a doctoral student of Dr. Nair, Deputy Director, National Institute of Cholera and Enteric Diseases. Her research interest is the epidemiology of enteric pathogens.
Acknowledgment
The work was supported, in part, by the Japan International Cooperation Agency (JICA/NICED Project 054-1061-E-O) and by a grant-in-aid for Scientific Research from the Ministry of Education, Science, Sports and Culture, Japan.
References
- Pal SC, Sircar BK, Nair GB, Deb BC. Epidemiology of bacterial diarrheal diseases in India with special reference to V. parahaemolyticus infections. In: Takeda Y, Miwatani T, editors. Bacterial diarrheal diseases. Tokyo: KTK Scientific Publishers; 1985: p. 65-73.
- Nishibuchi M, Kaper JB. Thermostable direct hemolysin gene of Vibrio parahaemolyticus: a virulence gene acquired by a marine bacterium. Infect Immun. 1995;64:2093–9.PubMedGoogle Scholar
- Sakurai J, Matsuzaki A, Miwatani T. Purification and characterization of thermostable direct hemolysin of Vibrio parahaemolyticus. Infect Immun. 1973;8:775–80.PubMedGoogle Scholar
- Honda T, Ni Y, Miwatani T. Purification and characterization of a hemolysin produced by a clinical isolate of Kanagawa phenomenon-negative Vibrio parahaemolyticus and related to the thermostable direct hemolysin. Infect Immun. 1988;56:961–5.PubMedGoogle Scholar
- Shirai H, Ito H, Hirayama T, Nakamoto Y, Nakabayashi N, Kumagai K, Molecular epidemiologic evidence for association of thermostable direct hemolysin (TDH) and TDH-related hemolysin of Vibrio parahaemolyticus with gastroenteritis. Infect Immun. 1990;58:3568–73.PubMedGoogle Scholar
- Suthienkul O, Ishibashi M, Tida T, Nettip N, Supavej S, Eampokalap B, Urease production correlates with possession of the trh gene in Vibrio parahaemolyticus strains isolated in Thailand. J Infect Dis. 1995;172:1405–8.PubMedGoogle Scholar
- Okuda J, Ishibashi M, Hayakawa E, Nishino T, Takeda Y, Mukhopadhyay AK, Emergence of a unique O3:K6 clone of Vibrio parahaemolyticus in Calcutta, India, and isolation of strains from the same clonal group from southeast Asian travellers arriving in Japan. J Clin Microbiol. 1997;35:3150–5.PubMedGoogle Scholar
- Bag PK, Nandi S, Bhadra RK, Ramamurthy T, Bhattacharya SK, Nishibuchi M, Clonal diversity among the recently emerged strains of Vibrio parahaemolyticus O3:K6 associated with pandemic spread. J Clin Microbiol. 1999;37:2354–7.PubMedGoogle Scholar
- World Health Organization. Vibrio parahaemolyticus, Japan, 1996-1998. Wkly Epidemiol Rec. 1999;74:361–3.PubMedGoogle Scholar
- Matsumoto C, Okuda J, Ishibashi M, Iwanaga M, Garg P, Ramamurthy T, Pandemic spread of an O3:K6 clone of Vibrio parahaemolyticus and emergence of related strains evidenced by arbitrarily primed PCR and toxRS sequence analyses. J Clin Microbiol. 2000;38:578–85.PubMedGoogle Scholar
- Wechsler E, D'Auo C, Mill VA, Mopper J, Myers-Wiley D, O'Keeffe E, Outbreak of Vibrio parahaemolyticus infection associated with eating raw oysters and clams harvested from Long Island Sound-Connecticut, New Jersey and New York, 1998. MMWR Morb Mortal Wkly Rep. 1999;48:48–51.PubMedGoogle Scholar
- Roy Chowdhury N, Chakraborty S, Eampokalap B, Chaicumpa W, Chongsa-Nguan M, Clonal dissemination of Vibrio parahaemolyticus displaying similar DNA fingerprint but belonging to two different serovars (O3:K6 and O4:K68) in Thailand and India. Epidemiol Infection. In press.
- Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980;8:4321–5. DOIPubMedGoogle Scholar
- Brosius J, Ullrich A, Raker MA, Gray A, Dull TJ, Gutell RR, Construction and fine mapping of recombinant plasmids containing the rrnB ribosomal RNA operon of E. coli. Plasmid. 1981;6:112–8. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 6, Number 6—December 2000
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
G. Balakrish Nair, National Institute of Cholera and Enteric Diseases, P-33, CIT Road, Scheme XM, Beliaghata, Calcutta - 700 010, India; fax: 91-33-3505066
Top