Volume 7, Number 4—August 2001
THEME ISSUE
West Nile Virus
West Nile Virus
West Nile Virus Infection in Birds and Mosquitoes, New York State, 2000
Abstract
West Nile (WN) virus was found throughout New York State in 2000, with the epicenter in New York City and surrounding counties. We tested 3,403 dead birds and 9,954 mosquito pools for WN virus during the transmission season. Sixty-three avian species, representing 30 families and 14 orders, tested positive for WN virus. The highest proportion of dead birds that tested positive for WN virus was in American Crows in the epicenter (67% positive, n=907). Eight mosquito species, representing four genera, were positive for WN virus. The minimum infection rate per 1,000 mosquitoes (MIR) was highest for Culex pipiens in the epicenter: 3.53 for the entire season and 7.49 for the peak week of August 13. Staten Island had the highest MIR (11.42 for Cx. pipiens), which was associated with the highest proportion of dead American Crows that tested positive for WN virus (92%, n=48) and the highest number of human cases (n=10).
The emergence of West Nile (WN) virus in 1999 in four U.S. states (1) was followed by its spread to 12 states in 2000 (2). An enzootic cycle was established between birds and mosquitoes, and WN disease was observed in humans, horses, and birds in both years (2,3). Bird deaths due to WN virus are unusual outside North America, with the exception of deaths of geese in Israel (4) and pigeons in Egypt (5). In 1999 in North America, WN disease, characterized by meningoencephalitis and myocarditis, was observed in 14 species of wild and captive birds (6). WN virus has been detected in a number of mosquito genera in North America, including Culex and Aedes species (2,7). Vector competence has been confirmed experimentally for some North American species, including Cx. pipiens, Ae. vexans, and Ae. japonicus (8,9).
We have summarized surveillance data for WN virus in dead birds and mosquitoes for New York State in the 2000 transmission season. A quantitative and kinetic analysis of data within and outside the epicenter is shown for both the bird and mosquito samples. Vertebrate and invertebrate WN virus infections are compared for counties in the epicenter.
Bird and Mosquito Samples
Dead birds were collected and mosquitoes were trapped by local county health units and other agencies as part of the New York State WN virus surveillance effort. Dead birds were necropsied at the Wildlife Pathology Unit at the Department of Environmental Conservation. Kidney, brain, heart, liver, or spleen were harvested and stored at -70°C. Additional avian tissue samples sent to the National Wildlife Health Center were not included in this analysis because selection criteria and testing procedures differed. Mosquitoes were trapped, speciated, grouped into pools of 5 to 50, and stored at -70°C. For some pools, Cx. pipiens and Cx. restuans were not separated but were pooled together and denoted as Cx. pipiens-restuans. The Aedes genus is being reclassified into two genera, Aedes and Ochlerotatus (10), but is classified as Aedes in this manuscript. Avian tissue samples and mosquito pools were sent to the Arbovirus Laboratory at the Wadsworth Center for WN virus testing. The transmission season was defined as May 15 to October 31, 2000, with the first and last positive samples collected on May 22 and October 31, 2000, respectively. Two positive birds were found earlier in the year (a hawk on February 6 and a crow on April 1), but they were not followed by other positive specimens and did not therefore appear to represent the beginning of the mosquito-borne WN virus transmission season.
WN Virus Testing
Samples were processed and WN virus assays were performed as described by Shi et al. (11). Briefly, RNA was extracted from bird tissue or pools of <50 mosquitoes, and real-time reverse transcription-polymerase chain reaction (RT-PCR) was performed (TaqMan, ABI Prism 7700 Sequence Detector, Applied Biosystems, Foster City, CA). Confirmatory tests included a second TaqMan primer-probe set, standard RT-PCR, virus isolation in cell culture, and immunofluorescence assays (for avian tissues). A sample was confirmed as positive if at least two different test results were positive.
Other Arbovirus Testing
Virus isolation was attempted for all Aedes species (with the exception of some Ae. vexans pools), all Culiseta species, and most WN virus-positive mosquito pools (with the exception of some Cx. pipiens and Cx. pipiens-restuans pools). Samples were inoculated onto monolayers of Vero or C6/36 cells. Viruses from positive cultures were typed by using serogroup-specific polyclonal antisera for bunyaviruses (California and Bunyamwera serogroups), flaviviruses, alphaviruses, and rhabdoviruses (Hart Park serogroup). California serogroup isolates were further characterized by sequence analysis.
Definition of the Epicenter and Radial Regions
The epicenter of the New York State epizootic was defined as follows. The minimum infection rate (MIR) of each mosquito species was calculated for each county or New York City borough by the standard formula: (number of WN virus-positive mosquito pools/total number of mosquitoes tested) x 1000. The MIR was calculated only when at least 1,000 mosquitoes were tested per species per county or borough. When the MIR was at least 1.0 for any mosquito species in a region, the county or borough was included in the epicenter. In addition, any counties that bordered at least two other epicenter counties or boroughs were included in the definition. The epicenter included the five boroughs of New York City (the Bronx, Brooklyn, Manhattan, Queens, and Staten Island) and the four counties immediately east and north of New York City (Nassau, Suffolk, Rockland, and Westchester counties). For both 1999 and 2000, all human cases of WN virus in New York State were in one of these counties (2,7).
Figure 1
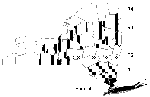
Figure 1. . Map of New York State showing the epicenter and radial regions used for analysis. The non-epicenter was defined as R1, R2, R3, and R4. Counties included in the regions are...
Counties outside the epicenter (the "non-epicenter") were divided into four radial regions, R1 to R4, with increasing distance from the epicenter (Figure 1). Radial regions were defined as follows: R1 = Putnam, Orange, Dutchess, Sullivan, and Ulster counties; R2 = Columbia, Delaware, Greene, Rensselaer, Montgomery, Albany, Otsego, Broome, Cortland, Schenectady, Schoharie, and Chenango counties; R3 = Fulton, Essex, Hamilton, Herkimer, Allegany, Lewis, Chemung, Madison, Cayuga, Schuyler, Yates, Washington, Warren, Tompkins, Tioga, Steuben, Onondaga, Seneca, Saratoga, Ontario, Oswego, and Oneida counties; and R4 = Monroe, Wyoming, Cattaraugus, Wayne, Chautauqua, Erie, Clinton, Genesee, Jefferson, Orleans, St. Lawrence, Niagara, Franklin, and Livingston counties.
During the 2000 transmission season, WN virus testing was performed on 3,403 dead birds, representing 153 species, 46 families, and 18 orders. The 1,201 WN virus-positive birds represented 63 species, 30 families, and 14 orders (Table 1). The percentage of WN virus-positive birds was 35% for all birds submitted for testing from throughout the state. Avian species that were >35% positive and for which at least 10 birds were tested included American Kestrel (57%, n=14), Cedar Waxwing (60%, n=10), Ovenbird (50%, n=18), American Crow (47%, n=1,687), Fish Crow (47%, n=45), and Red-tailed Hawk (43%, n=14). The discrepancies in number of birds tested make comparisons between species difficult.
The percentage of WN virus-positive birds was analyzed for the epicenter and non-epicenter regions. Data are included for avian species for which at least 10 birds were tested in one of the regions (Table 2). For all submitted birds, 51% and 23% WN virus-positive birds were found in the epicenter and non-epicenter regions, respectively. WN virus infection in dead birds was highest for American Crows (67%) in the epicenter. In the non-epicenter, WN virus infection for crows was lower, similar to infection in all birds in this region. High numbers of crows were tested in both regions, and the percentage positive differed by almost threefold.
Figure 2
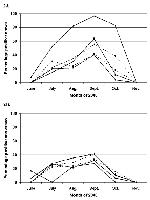
Figure 2. . Percentage of West Nile virus-positive birds in different regions of New York State over time. Percentage of positive American Crows (2A) or birds excluding American Crows (2B) in the epicenter...
WN virus infection in dead birds was examined over time for American Crows and all other birds in the epicenter and four radial regions in New York State (Figure 2). Comparison of American Crows in the five regions (Figure 2A) revealed the highest peak in the epicenter during September. In addition, the peak for American Crows in the epicenter was much broader than for the other four regions. For September, the peaks for American Crows in R1 and R2 were greater than those in the more distant regions, R3 and R4, suggesting a minor extension of the epicenter during this month. Comparison of all birds except American Crows in the five regions (Figure 2B) revealed little difference between the regions, even for the epicenter. These data support the hypothesis that the susceptibility to WN disease was greatest in crows in the epicenter.
Figure 3
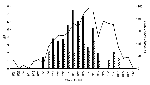
Figure 3. . Weekly minimal infection rate per 1,000 mosquitoes (MIR) and percentage of crows positive for West Nile virus in the epicenter. Solid bars designate Culex pipiens. Hatched bars designate Cx. pipiens-restuans....
We tested 9,954 mosquito pools with 317,668 mosquitoes, representing 28 species and eight genera (Table 3). Of eight positive species representing four genera, most positive pools were Culex species (n=341), compared with only 22 positive pools in the other three genera. All but five of the positive pools were collected in the epicenter. The MIR was calculated for each species in the epicenter for which at least 1,000 mosquitoes were tested (Table 3). In the epicenter, the MIR ranged from 0.47 to 3.55. The MIR of Cx. pipiens was the highest for an individual species. All the pure Cx. restuans pools were negative, and the MIR for Cx. pipiens-restuans was almost half that of the pure Cx. pipiens pools; therefore, the positive mosquitoes in the mixed pools of Cx. pipiens-restuans were most likely Cx. pipiens. The MIRs for Cx. pipiens and Cx. pipiens-restuans were compared with the percentage of positive American Crows in the epicenter for each week of the transmission season (Figure 3). The MIRs for Cx. pipiens and Cx. pipiens-restuans peaked 3 and 2 weeks, respectively, before the peak for positive crows. These data support the hypothesis that Cx. pipiens is an important enzootic vector of WN virus in New York.
The epicenter was examined as individual boroughs and counties to compare vertebrate and invertebrate WN virus infections (Table 4). The percentage of positive American Crows was calculated, and human and equine cases were noted for each county over the entire season. The MIRs of mosquitoes from each county were calculated for species with at least 1000 mosquitoes tested. Six mosquito species or groups met this criterion. The highest number of vertebrates infected with WN virus was found in Staten Island and was associated with the highest mosquito MIRs. This borough had measurable MIRs for five mosquito groups: Cx. pipiens, Cx. species, Cx. pipiens-restuans, Cx. salinarius, and Ae. vexans.
Virus isolation was performed on mosquito samples by cell culture. WN virus was isolated from 110 samples, including Cx. pipiens-restuans (n=60), Cx. pipiens (n=25), Cx. salinarius (n=13), Culex species (n=9), Ae. triseriatus (n=1), Ae. vexans (n=1), and Psorophora ferox (n=1). No virus was isolated from any of the WN virus RNA-positive pools of Ae. japonicus, Ae. cantator, and An. punctipennis. Pools that were negative on Vero cell culture were passaged repeatedly in Aedes albopictus (C6/36) cells; these further attempts to isolate virus were unsuccessful. Other viruses isolated were trivittatus virus from Ae. trivittatus (n=4) and Ae. triseriatus (n=1), Cache Valley virus from Ae. trivittatus (n=2) and Ae. triseriatus (n=2), and Flanders virus from Cx. pipiens-restuans (n=7), Cx. pipiens (n=2), and Cs. melanura (n=11). Eastern equine encephalitis virus and California group viruses other than trivittatus were not isolated from any of the Culiseta or Aedes pools.
In the 2000 transmission season in New York State, we found 63 bird species infected with WN virus, compared with 14 species in 1999 (6). The percentage of WN virus-positive birds was higher in the epicenter than outside it. This high percentage almost entirely reflects infected crows in the epicenter; no increase in WN virus infection was noted in birds other than crows. In contrast, high WN virus infection of dead crows was not observed outside the epicenter, where the percentage of WN virus positivity was similar in crows and other birds over the entire season. High numbers of dead crows were also observed in 1999 (3,6). The cause of the increased sensitivity of crows to WN disease or infection is unknown, but may be due to virus-host interactions, mosquito-bird interactions, mosquito feeding preferences, crow population density, or crow behavior. The presence of WN virus in dead birds does not indicate a definitive diagnosis of WN virus as the cause of death. Many of the birds did not show gross pathologic lesions of WN disease (12). In addition, the rate of WN virus-positive birds in our surveillance samples is not equivalent to prevalence of infection, since we are sampling only dead birds.
The analysis of percentage positive birds over time revealed differences between various regions in New York State. The percentage of WN virus-positive crows was highest in the epicenter compared with other regions of New York State throughout the season, supporting the importance of crows as indicators in the epizootic. The percentage of WN virus-positive crows was higher than that for all other birds early in the season only in the epicenter. Outside the epicenter, the percentages of WN virus-positive crows and all other birds were similar until the peak month of September. At this time, the two radial regions closest to the epicenter showed higher infection in crows than in other birds, suggesting that the intense level of viral activity may have spread beyond the epicenter. An explanation for this apparent spread may be increased movement of crows during the fall. The surveillance data on avian deaths have implications for future surveillance activities. The similar percentages of positive crows and other birds outside the epicenter indicate the importance of testing all birds, not only crows, outside the epicenter.
Sampling errors are likely with the avian surveillance samples. The samples were from dead birds submitted to the Wildlife Pathology Unit, which relied on the cooperation of the general public, individual county health departments, and other agencies; therefore, surveillance samples do not represent a random sampling of dead birds throughout the state. The size and degree of urbanization of various bird species may have resulted in differences in submission. For example, large urban-dwelling species, such as crows, were more likely to have been submitted than small rural dwellers. In addition, more birds were likely sampled from areas with human cases, greater media coverage, and higher human population. The similarity in WN virus infection in birds other than crows from different regions suggests that the impact of sampling bias was not significant. Additionally, specimens submitted for testing to the Wadsworth Center from the Wildlife Pathology Unit represent a sample of those submitted by the public. Sampling at this level may introduce further bias into our surveillance sample. The similarity in WN virus infection in birds other than crows outside the epicenter, however, suggests that such bias was minimal.
The highest MIR for mosquitoes in the epicenter was for Cx. pipiens. Positive pools of Cx. pipiens also were identified in 1999 in New York (13). Cx. pipiens-pipiens mosquitoes feed almost exclusively on birds (14); thus, they are likely an important enzootic vector in the bird-mosquito cycle in North America. Other Cx. species have been implicated as enzootic vectors worldwide (15). High MIRs of Cx. pipiens and Cx. salinarius were associated with human and equine WN virus cases and high infection rates of crows in counties in the epicenter. Cx. salinarius feeds on both birds and mammals (16); therefore, it is a likely candidate as a "bridge vector," transmitting the virus from bird to mammal. Cx. salinarius has been proposed as a bridge vector for Eastern equine encephalitis virus (17). All MIR data were calculated by using RT-PCR-positive pools and therefore cannot be directly compared with MIRs calculated by using infectious virus-positive pools. No virus was isolated from five RNA-positive pools of Ae. japonicus, even after six serial passages through C6/36 cells. Much attention has been focused on this species because it has been reported to be a highly competent laboratory vector of WN virus (9), but current field data do not support this experimental observation. Different populations of Ae. japonicus have been described in the eastern United States (18), and differences in vector competence between the populations may explain the discrepancy between the field and experimental data.
The possibility of sampling error also exists for the mosquito surveillance samples. Individual counties collected mosquitoes in different numbers and set traps by different criteria. In addition, some counties used mosquito adulticides or larvicides during the season. These sources of bias are unlikely to have been uniformly introduced, and their impact on our analyses is unclear.
The results from the 2000 surveillance season for WN virus leave a number of unanswered questions. Many avian species can become infected with WN virus, but the prevalence of infection for each species is unknown without systematic serosurveys of the wild bird population. It is also unknown which birds have a high enough viremia for efficient transmission to the vector. The apparent mortality rate caused by WN virus is higher for crows than for other birds, but laboratory experiments are required to determine WN virus mortality rates and the pathogenic mechanisms in crows and other avian species. In addition to crows, many other birds, such as raptors and other corvids, also showed significant pathology. Some nonmigrating species (e.g., ruffed grouse) have potential use as an indicator species for WN virus infection. Vector competence, blood meal identification, and transovarial transmission studies for the potential mosquito vectors are also important research areas.
Dr. Bernard is assistant director of the Arbovirus Laboratory at the Wadsworth Center. Her major research interests are viral pathogenesis and the maintenance of arboviruses in temperate climates.
Acknowledgments
We thank the WN virus surveillance team, especially the New York City Department of Health and the local county health units, who collected the surveillance specimens. We also thank H. Taber for helpful discussions.
Support was provided by the Wadsworth Center and the Division of Epidemiology, New York State Department of Health, the Department of Environmental Conservation, and the Centers for Disease Control and Prevention.
References
- Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science. 1999;286:2333–7. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Update: West Nile Virus activity--Eastern United States, 2000. MMWR Morb Mortal Wkly Rep. 2000;49:1044–7.PubMedGoogle Scholar
- Centers for Disease Control and Prevention. Outbreak of West Nile-like viral encephalitis--New York, 1999. MMWR Morb Mortal Wkly Rep. 1999;48:845–9.PubMedGoogle Scholar
- Office of Internationale des Epizooties. West Nile fever in Israel in geese. Disease Information. 1999;12:166.
- Work TH, Hurlbut HS, Taylor RM. Isolation of West Nile virus from hooded crow and rock pigeon in the Nile Delta. Proc Soc Exp Biol Med. 1953;84:719–22.PubMedGoogle Scholar
- Steele KE, Linn MJ, Schoepp RJ, Komar N, Geisbert TW, Manduca RM, Pathology of fatal West Nile virus infections in native and exotic birds during the 1999 outbreak in New York City, New York. Vet Pathol. 2000;37:208–24. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Update: West Nile virus encephalitis--New York, 1999. MMWR Morb Mortal Wkly Rep. 1999;48:944–55.
- Turell MJ, O'Guinn M, Oliver J. Potential for New York mosquitoes to transmit West Nile virus. Am J Trop Med Hyg. 2000;62:413–4.PubMedGoogle Scholar
- O'Guinn ML, Dohm DJ, Jones JW, Turell MJ. Potential mosquito vectors of West Nile virus in the United States. Am J Trop Med Hyg. 2001;62:286.
- Reinert JF. New classification for the composite genus Aedes (Diptera: Culicidae: Aedini), elevation of subgenus Ochlerotatus to generic rank, reclassification of other subgenera, and notes on certain subgenera and species. J Am Mosq Control Assoc. 2000;16:175–88.PubMedGoogle Scholar
- Shi P-Y, Kauffman EB, Ren P, Felton A, Tai JH, Dupuis AP II, High throughput detection of West Nile virus RNA. J Clin Microbiol. 2001;39:1264–71. DOIPubMedGoogle Scholar
- Eidson M, Kramer LD, Stone W, Hagiwara Y, Schmit K; New York State West Nile Virus Avian Surveillance Team. Dead bird surveillance as an early warning system for West Nile virus. Emerg Infect Dis. 2001;7:631-5.PubMedGoogle Scholar
- Centers for Disease Control and Prevention. Update: West Nile-like viral encephalitis--New York, 1999. MMWR Morb Mortal Wkly Rep. 1999;48:890–2.PubMedGoogle Scholar
- Spielman A. Population structure in the Culex pipiens complex of mosquitos. Bull World Health Organ. 1967;37:271–6.PubMedGoogle Scholar
- Hubalek Z, Halouzka J. West Nile fever--a reemerging mosquito-borne viral disease in Europe. Emerg Infect Dis. 1999;5:643–50. DOIPubMedGoogle Scholar
- Crans WJ. Continued host preference studies with New Jersey mosquitoes, 1963. In: Proceedings of the Annual Meeting of the New Jersey Mosquito Extermination Association 1964;51:50-8.
- Vaidyanathan R, Edman JD, Cooper LA, Scott TW. Vector competence of mosquitoes (Diptera: Culicidae) from Massachusetts for a sympatric isolate of eastern equine encephalomyelitis virus. J Med Entomol. 1997;34:346–52.PubMedGoogle Scholar
- Fonseca DM, Campbell SR, Crans WJ, Mogi M, Miyagi I, Toma T, Aedes (Finlaya) japonicus (Diptera: Culicidae), a newly recognized mosquito in the United States: analyses of genetic variation in the United States and putative source populations. J Med Entomol. 2001;38:135–46. DOIPubMedGoogle Scholar
Figures
Tables
Cite This Article1P. Bryon Backenson, Ivan Gotham, Yoichiro Hagiwara, Geraldine S. Johnson, Gary Lukacik, Kathryn Schmit, and Amy L. Willsey, Division of Epidemiology, New York State Department of Health
Table of Contents – Volume 7, Number 4—August 2001
| EID Search Options |
|---|
|
|
|
|
|
|