Volume 9, Number 10—October 2003
Dispatch
West Nile Virus Transmission in Resident Birds, Dominican Republic
Abstract
We report West Nile virus (WNV) activity in the Dominican Republic for the first time. Specific anti-WNV antibodies were detected in 5 (15%) of 33 resident birds sampled at one location in November 2002. One seropositive bird was <4 months old, indicating a recent infection.
The initial outbreak of West Nile virus (WNV; family Flaviviridae, genus Flavivirus) in the Western Hemisphere took place in New York in 1999, with deaths observed in humans, horses, and numerous species of wild birds (1). Since then, this virus has spread rapidly across North America (2,3). Migratory birds are suspected of being responsible for the rapid spread of WNV through North America (4), and transport of WNV by Neotropical migratory birds throughout the New World has been anticipated (5).
Although WNV has spread rapidly through continental areas, its ability to spread across oceanic barriers is uncertain. The many islands of the West Indies represent the wintering grounds of numerous North American migratory birds (6,7) that breed in or migrate through WNV transmission foci in the United States. The Caribbean islands tend to have high human population density, and low populations of many birds and other vertebrates are restricted to certain islands. Introduction of WNV to the West Indies would present a human and equine health concern and potentially threaten numerous endangered and endemic bird species and perhaps other wild vertebrates.
Given the speculation that WNV may be disseminated by migrating birds (5,8,9), we hypothesized that the virus would be introduced to the Dominican Republic. Accordingly, we sampled apparently healthy birds there for evidence of locally acquired WNV infection.
Figure
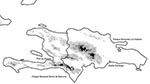
Figure. Hispaniola in the West Indies, on which are located Haiti (western third of the island) and the Dominican Republic. West Nile virus transmission occurred at Parque Nacional Los Haitises before November...
Birds were studied at two sites in the Dominican Republic, on the island of Hispaniola (Figure): Parque Nacional Sierra de Baoruco (November 7–16, 2002; 18° 12' N, 71° 32' W) and Parque Nacional Los Haitises (November 18–23, 2002; 19° 00' N, 69° 30' W). Birds were collected by standard methods (10). Tissues (eye, spleen, and kidney) were removed from 89 birds of 29 species (25 resident, 4 migratory) at Sierra de Baoruco and from 58 birds of 27 species (18 resident, 9 migratory) at Los Haitises; the tissues were tested for active WNV infection. Blood samples were collected from a subsample of these birds, including 41 that represented 18 resident species at Sierra de Baoruco and 33 that represented 16 resident species at Los Haitises. Blood was not collected from migratory birds or from certain very small resident birds, such as hummingbirds. Blood and tissue specimens were frozen immediately in liquid nitrogen for transportation and then stored at –70°C. Voucher specimens (including additional tissue samples) were prepared for all birds and are deposited at the University of Kansas Natural History Museum (KUNHM).
The sex and breeding condition of each bird were determined by examination and measurement of gonads. Birds’ ages were assessed by plumage, skull ossification, and presence or absence of a bursa of Fabricius. The migratory or resident status of each bird was determined on the basis of standard references (6,7). For species that had both migratory and resident populations, we based status assessment on breeding conditions (breeding birds were assumed to be resident).
Serum samples were screened for flavivirus-neutralizing antibodies by plaque-reduction neutralization test (PRNT) according to standard methods (11) and by using challenge inocula of 100 plaque-forming units (PFU) WNV strain NY99-4132 and Saint Louis encephalitis virus (SLEV) strain TBH-28. Testing for neutralizing antibody to SLEV was important because this virus has been detected in the Caribbean and isolated from wild birds (12) and cross-reacted to anti–WNV-neutralizing antibodies in 6% of seropositive birds sampled in New York (13). PRNTs were performed with Vero cells in 6-well plates and a serum dilution of 1:10 in BA1 buffer (Hanks M-199 salts, 0.05 M Tris pH 7.6, 1% bovine serum albumin, 0.35 g/L sodium bicarbonate, 100 U/mL penicillin, 100 mg/L streptomycin, 1 mg/L Fungizone). Specimens that neutralized the virus stocks by at least 80% were further titrated in duplicate. To identify either virus as the causative agent, we used 90% neutralization as the criterion for a positive test result, and a four fold greater titer to one of the flaviviruses was considered diagnostic for that flavivirus.
Serum samples that showed neutralizing antibody titers to WNV or SLEV were tested by epitope-blocking enzyme-linked immunosorbent assays (ELISAs) with the WNV-specific monoclonal antibody 3.1112G, which discriminates between WNV and SLEV infections in birds (14). Because of the current lack of information on flaviviruses in the Dominican Republic, we required both the PRNT and the ELISA to test positive to consider a serum sample positive for WNV.
Tissues from each individual bird were pooled and homogenized in 2 mL of BA1 supplemented with 20% fetal bovine serum. Homogenates were clarified by centrifugation at 3,700 rpm for 10 min at 4°C. Four hundred microliters of each homogenate was screened for virus by Vero plaque assay (11). Homogenates obtained from flavivirus-seropositive birds were assayed for WNV RNA by TaqMan reverse-transcription polymerase chain reaction with WNV-specific primers (15).
Serum samples from nine resident birds tested positive for flavivirus-neutralizing antibodies (Table). Of these, five birds were positive for WNV antibodies by PRNT and blocking ELISA. All WNV antibody–positive birds were sampled at the Los Haitises study site. One serum sample was collected from an immature bird (ruddy quail-dove, Geotrygon montana; KUNHM 94667) that was <4 months old, suggesting that virus transmission was recent. Virus was not isolated from any of the tissues tested from 118 resident and 29 migratory birds (all migrants were Parulidae), nor did we detect WNV RNA in any of the tissue homogenates from flavivirus-seropositive birds.
Our finding of WNV-neutralizing antibodies in five resident birds represents the first evidence of WNV activity in the Dominican Republic. No cases of WNV infection in humans, horses, or birds were known at the time of sampling. The birds in this study could have been infected with WNV in the Dominican Republic as recently as early November 2002; nonetheless, the virus probably arrived earlier in the Caribbean region. Because no current infections were detected, our results reflect past virus transmission activity. Although we cannot determine when this activity began, the seropositive immature quail-dove presumably was infected after mid-July 2002, when it was born, and before early November 2002, in order to have stimulated detectable antibody production by mid-November.
The earliest evidence of WNV transmission in the West Indies is a human case from the Cayman Islands in 2001 (16). WNV-seropositive birds captured in Jamaica early in 2002 may have been infected in 2001 or earlier (17). Additional evidence of WNV transmission in the Caribbean region includes the report of two seropositive horses in Yucatán state, Mexico, sampled in July 2002 (18). Although the seropositivity of vertebrates in Cayman Islands, Jamaica, Mexico, and now the Dominican Republic is strong evidence for WNV activity in the region, it is indirect evidence and does not entirely rule out the possibility of cross-reactions with another flavivirus in laboratory assays. WNV remains to be isolated from the region.
The presence of WNV at Los Haitises may have resulted from transportation by viremic migratory birds from North America, where WNV transmission foci are widespread (2). Several migratory bird species, in particular parulid warblers (order Passeriformes), were observed at this site. At least some passerine birds are capable of transmitting virus during their few days of viremia (19). Therefore, transmission to mosquitoes or predators from viremic migrants would be possible for a brief period (a few days, at most) after arrival at a site. Virus introduction into Caribbean ecosystems is therefore likely to occur at coastal sites where transoceanic migrants make first landfall.
We found no evidence of active virus in bird tissues of both resident and migratory species. We did not test serum samples from migrants because the presence of antibodies would not be informative, given the history of these birds traveling through areas of WNV transmission in or near North American breeding grounds. We presume that the five seropositive resident birds were infected locally because the four species involved are not migratory (7). Although young birds may disperse several kilometers from natal sites (20), adults probably live entirely within breeding territories.
Although only five birds (15%, 95% confidence interval [CI] 5% to 32%) at Los Haitises were found to be seropositive, the results suggest that transmission of WNV among bird populations at that site was widespread. For comparison, seroprevalence of WNV-neutralizing antibodies in resident birds was 50% (CI 44% to 57%) in Queens, New York City, after the 1999 outbreak (13), and 23% (CI 18% to 29%) in Staten Island after the 2000 outbreak (21). We used conservative criteria for determining a positive result because the background diversity of flaviviruses in the Dominican Republic has not been studied recently. If only the PRNT had been used (as was the case in the New York studies), then seven (21%, CI 7% to 35%) of the birds from Los Haitises would have been reported as positive for antibodies to WNV.
The evidence for local WNV transmission in the Dominican Republic indicates risk for West Nile fever and meningoencephalitis in the human, equine, and avian populations of Hispaniola. We suggest that WNV be considered in the differential diagnosis of humans and other vertebrates with central nervous system disease in Hispaniola.
Mr. Komar is an ornithologist working on a Ph.D. in biology at the University of Kansas Natural History Museum and Department of Ecology and Evolutionary Biology in Lawrence, Kansas. His primary research interests are avian ecology and conservation in the Neotropical region.
Acknowledgments
We thank Bolivar Cabrera, Tristan Davis, Juan Cepedes Hidalgo, José Dolores Jiménez, Esteban López-Medrano, and Domingo Sirí for assistance in the field; José Ramón Albaine and Simón Guerrero for logistical assistance and assistance with permits; John Roehrig for assisting with the interpretation of serologic test results; and the two anonymous reviewers who provided comments on the manuscript.
This study was supported in part by National Science Foundation Grant #0211388, and in part by the Centers for Disease Control and Prevention contract U50/CCU 820510-02.
References
- Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science. 1999;286:2333–7. DOIPubMedGoogle Scholar
- Chow CC, Montgomery SP, O’Leary DR, Nasci RS, Campbell GL, Kipp AM, Provisional surveillance summary of the West Nile virus epidemic—United States, January–November 2002. MMWR Morb Mortal Wkly Rep. 2002;51:1129–33.PubMedGoogle Scholar
- Health Canada. Population and Public Health Branch WNV surveillance updates, December 4, 2002. Available from: URL: http://www.hc-sc.gc.ca/pphb-dgspsp/wnv-vwn/mon_e.html#sitrep
- McLean RG, Ubico SR, Bourne D, Komar N. West Nile virus in livestock and wildlife. Curr Top Microbiol Immunol. 2002;267:271–308.PubMedGoogle Scholar
- Rappole JH, Derrickson SR, Hubalek Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg Infect Dis. 2000;6:319–28. DOIPubMedGoogle Scholar
- American Ornithologists’ Union. Check-list of North American birds. 7th ed. Washington: American Ornithologists’ Union; 1998.
- Raffaele H, Wiley J, Garrido O, Keith A, Raffaele J. A guide to the birds of the West Indies. Princeton (NJ): Princeton Univ. Press; 1998.
- Malkinson M, Banet C, Weisman Y, Pokamunski S, King R, Drouet MT, Introduction of West Nile virus in the Middle East by migrating white storks. Emerg Infect Dis. 2002;8:392–7. DOIPubMedGoogle Scholar
- Peterson AT, Vieglais DA, Andreasen JK. Migratory birds modeled as critical transport agents for West Nile virus in North America. Vector Borne Zoonotic Dis. 2003;3:27–37. DOIPubMedGoogle Scholar
- Gaunt AS, Oring LW, eds. Guidelines to the use of wild birds in research. The Ornithological Council; 1997. Available from: URL: www.nmnh.si.edu/BIRDNET/GuideToUse
- Beaty BJ, Calisher CH, Shope RE. Arboviruses. In: Lennette EH, Lennette DA, Lennette ET, editors. Diagnostic procedures for viral, rickettsial, and chlamydial infections, 7th ed. Washington: American Public Health Association; 1995. p. 189–212.
- Tsai TF, Mitchell CJS. Louis encephalitis. In: Monath TP, editor. Vol. IV, The arboviruses: epidemiology and ecology. Boca Raton (FL): CRC Press; 1988. p. 113–43.
- Komar N, Panella NA, Burns JE, Dusza SW, Mascarenhas TM, Talbot TO. Serologic evidence for West Nile virus infection in birds in the New York City vicinity during an outbreak in 1999. Emerg Infect Dis. 2001;7:621–5. DOIPubMedGoogle Scholar
- Blitvich BJ, Marlenee NL, Hall RA, Calisher CH, Bowen RA, Roehrig JT, Epitope-blocking enzyme-linked immunosorbent assays for the detection of serum antibodies to West Nile virus in multiple avian species. J Clin Microbiol. 2003;41:1041–7. DOIPubMedGoogle Scholar
- Lanciotti RS, Kerst AJ, Nasci RS, Godsey MS, Mitchell CJ, Savage HM, Rapid detection of West Nile virus from human clinical specimens, field-collected mosquitoes, and avian samples by a TaqMan reverse transcriptase-PCR assay. J Clin Microbiol. 2000;38:4066–71.PubMedGoogle Scholar
- O’Leary DR, Nasci RS, Campbell GL, Marfin AA. West Nile virus activity—United States, 2001. MMWR Morb Mortal Wkly Rep. 2002;51:497–501.PubMedGoogle Scholar
- Dupuis AP II, Marra PP, Kramer LD. Serologic evidence for West Nile virus transmission in Jamaica, West Indies. Emerg Infect Dis. 2003;9:860–3.PubMedGoogle Scholar
- Loroño-Pino MA, Blitvich BJ, Farfán-Ale JA, Puerto FI, Blanco JM, Marlenee NL, Serologic evidence of West Nile virus infection in horses, Yucatan State, Mexico. Emerg Infect Dis. 2003;9:857–9.PubMedGoogle Scholar
- Komar N, Langevin S, Hinten S, Nemeth N, Edwards E, Hettler D, Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg Infect Dis. 2003;9:311–22.PubMedGoogle Scholar
- Greenwood PJ. Mating systems, philopatry and dispersal in birds and mammals. Anim Behav. 1980;28:1140–62. DOIGoogle Scholar
- Komar N, Burns J, Dean C, Panella NA, Dusza S, Cherry B. Serologic evidence for West Nile virus infection in birds in Staten Island, New York, after an outbreak in 2000. Vector Borne Zoonotic Dis. 2001;1:191–6. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 9, Number 10—October 2003
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Oliver Komar, Natural History Museum and Biodiversity Research Center, University of Kansas, 1345 Jayhawk Boulevard, Lawrence, KS 66045-7561, USA; fax: 785-864-5335
Top