Volume 29, Number 9—September 2023
Dispatch
Novel Echarate Virus Variant Isolated from Patient with Febrile Illness, Chanchamayo, Peru
Abstract
A new phlebovirus variant was isolated from an acute febrile patient in Chanchamayo, Peru. Genome characterization and p-distance analyses based on complete open reading frames revealed that the virus is probably a natural reassortant of the Echarate virus (large and small segments) with a yet-unidentified phlebovirus (M segment).
The genus Phlebovirus (order Bunyavirales, family Phenuiviridae) consists of 66 species according to the International Committee on Taxonomy of Viruses (1). Phleboviruses are globally distributed and can be transmitted by phlebotomine sandflies, mosquitoes, or ticks (2,3). Sandfly phlebovirus can cause unspecific symptoms in humans and often is misdiagnosed as dengue fever, malaria, or influenza (4,5); however, its clinical symptoms can range from high fever, severe headache, muscle pain, and aseptic meningitis to mild or severe meningoencephalitis (6). In Peru, 3 of 9 phleboviruses that cause febrile illness in Central and South America (3–5,7) have been identified: Echarate virus (ECHV), Maldonado virus (7), and Candiru virus (7).
During the last decade, isolates characterized by whole-genome sequencing have contributed to increased detection of novel and recombinant pathogenic and nonpathogenic phleboviruses worldwide (2,5,7), demonstrating a high viral diversity within this genus. Therefore, continuous public health surveillance, including genome characterization as a complementary tool, is critical to identifying novel and emerging viruses of clinical relevance in the Americas. We report the identification and characterization of a novel ECHV virus variant isolated from a patient with acute febrile illness (AFI) in Peru.
Figure 1
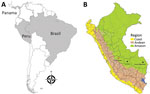
Figure 1. Geographic distribution of Candiru complex virus in Central and South America in study of novel ECHV variant isolated from patient with febrile illness, Chanchamayo, Peru. A) Countries where viruses were...
As part of passive clinic-based surveillance for AFI in Peru approved by Peru’s Ministry of Health and the US Naval Medical Research Unit South Institutional Review Board (protocol no. NMRCD.2010.0010) (8), a 20-year-old man who worked in civil construction was admitted to Hospital Regional Docente de Medicina Tropical Julio César Demarini Caro, located in the city of Chanchamayo in the northern region of Junín Department in central Peru, on June 25, 2019 (Figure 1). He had a 2-day history of fever, malaise, chills, systemic muscle pain, arthralgias, generalized head pain, drowsiness, photophobia, retroocular pain, and anorexia. He had conjunctival injection and an axillary temperature of 39.0°C, and the tourniquet test was negative.
We inoculated the acute serum sample into African green monkey kidney cells (Vero ATCC CCL-81) and Aedes albopictus mosquito cells (C6/36) and maintained them at 37°C (Vero cells) and 33°C (mosquito cells). The sample showed ≈50% cytopathic effect at day 7 in Vero cells, but no cytopathic effect occurred in C6/36 cells after 10 days. We prepared spot slides of both cell lines and tested them by indirect immunofluorescence assay (IFA) using pooled polyclonal antisera against flaviviruses (yellow fever virus and dengue virus serotype 3), alphaviruses (Mayaro virus and Venezuelan equine encephalitis virus), orthobunyaviruses (Oropouche virus, Guaroa virus, Caraparu virus, and Maguari virus), arenaviruses (Allpahuayo virus and Tacaribe virus), and cardiovirus. Only the Vero cells spot slide was reactive by IFA (≈25% of cells fluoresced) with pooled bunyaviruses polyclonal antibody. The second IFA with individual polyclonal antibody components also detected a weak reaction (≈25% of cells fluoresced) with Oropouche and Maguari polyclonal antibodies. Because of a weak positive signal at this level, we submitted the isolate for molecular characterization.
We extracted RNA from infected Vero cell supernatant by using the QIAamp Viral RNA Mini Kit (QIAGEN, https://www.qiagen.com), according to the manufacturer’s instructions. We amplified the viral genome by using 2 unbiased approaches, a modified sequence-independent, single-primer amplification (SISPA) protocol (9), and whole-transcriptome amplification (WTA) (10) using REPLI-g WTA Single Cell Kit (QIAGEN) according to manufacturer’s guidelines. We prepared libraries by using Nextera XT DNA Library Preparation Kit (Illumina, https://www.illumina.com) and sequenced them on the Illumina MiSeq platform by using MiSeq Reagent Kit version 3 (600-cycle) according to the manufacturer’s instruction.
We processed raw reads from both sequencing approaches (SISPA and WTA) for quality control, de novo assembly, host read subtraction, taxonomic classification, and gene family analysis by using 3 different bioinformatics pipelines: EDGE Bioinformatics tools (11), VirusSeeker (12), and MetaDetector (K.A. Bishop-Lilly et al., unpub. data). Both unbiased methods showed similar read quality. Results of taxonomic analysis of the reads and contigs obtained from SISPA and WTA showed Candiru phlebovirus as the unique human viral pathogen in the isolate, indicating that both techniques successfully amplified the isolated virus. We searched consensus sequences (GenBank accession nos. OQ623470–2) against a nucleotide database by using BLASTn and protein database by using BLASTx (both at https://blast.ncbi.nlm.nih.gov). Candiru phlebovirus large and small segments had >95% amino acid identity compared with those of ECHV. Of note, the medium (M) segment had 76.5% identity with that of ECHV at nucleotide level and 86.36% identity at amino acid level (Table 1). The M segment typically encodes for 3 polypeptides (NSm, Gn, and Gc), which are co-translationally cleaved. The NSm polypeptide is a virulence factor associated with the inhibition of apoptosis in infected cells and plays a role in viral mosquito infection (13). The amino acid identity value of the predicted NSm sequence of our isolated virus ranged from <30% with the other members of Candiru complex to 78.6% with ECHV. For average coverage calculation, we mapped the trimmed reads back to contigs obtained from de novo assembly and to the corresponding Echarate reference sequences as an orthogonal verification at 0.8 length fraction and 0.8 similarity fraction. The minimum coverage was 1,583× in the M segment with ECHV and the maximum was 10,224× in the M segment with the obtained contig (Table 2).
We performed pairwise sequence comparison between our isolate and Candiru complex viruses for the RNA-dependent RNA polymerase, glycoprotein precursor, nucleoprotein, and nonstructural genes. The p-distance value of the glycoprotein precursor gene 0.237 (nucleotide) between our isolate and ESCV was similar to those observed among other members of the Candiru complex (Appendix 1 Table) and consistent with values previously reported in the literature among members of the Candiru complex (0.2–0.46) and among members of the same complex but not among strains of the same virus (0.01–0.12) (7,14,15). Furthermore, considering the new variant was isolated and characterized 21 years later, we also calculated the overall mean distance values (0.079 for nucleotide, 0.039 for amino acid) for 74 complete M segment sequences of Oropouche virus published over time (1955–2021 [67 years]). Those values suggest that the difference could not be explained by virus mutation because the new isolate has a nucleotide difference value of 0.24 with ECHV. The low M segment identity value together with distance values probably indicate the uniqueness of this segment and support the concept that this is a novel ECHV variant that could be generated by a recombinant event between ECHV and an unknown phlebovirus.
Figure 2
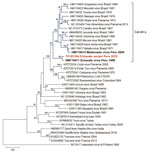
Figure 2. Maximum-likelihood phylogenetic tree based on 36 amino acid sequences of phleboviruses M segment (NSm–Gn) in study of novel Echarate virus variant isolated from patient with febrile illness, Chanchamayo, Peru. Strains...
To determine the evolutionary relationship of our isolate to other known members of the genus, we conducted maximum-likelihood phylogenetic analyses on the aligned amino acid sequence of the RNA-dependent RNA polymerase, glycoprotein precursor, nucleoprotein, and nonstructural genes (Appendix 2). All the phylogenetic trees placed our isolate among the Candiru virus complex within a well-supported clade with ECHV. However, the NSm or glycoprotein tree clustered the new variant together with ECHV within a well-supported clade separate from other Candiru complex viruses (Figure 2).
Our findings indicate that a novel ECHV variant is circulating in the jungle of central Peru. Because the clinical symptoms of infection with this variant are also characteristic of dengue, malaria, and other tropical infectious diseases common in this region (4,5) continued AFI biosurveillance is needed to detect novel and emerging pathogens to protect the health of the population and US service members deployed in affected areas in Peru. Ecologic studies are necessary to determine how widespread the new variant is within this region, to identify potential vectors and reservoirs involved in its transmission, and to support decision-making for keeping service members medically prepared and protected from health and safety threats both on and off duty.
Ms. Troncos is a research technician in the Virology and Emerging Infections Department of US Naval Medical Research Unit SOUTH, Lima, Peru. Her primary research interests include identification and characterization of viruses that cause illnesses in humans.
Acknowledgments
We thank the local Ministry of Health authorities in Peru, Red de Salud Chanchamayo, and Hospital Docente de Medicina Tropical Julio César Demarini Caro for their support to allow us to execute this study. We thank Patricia Aguilar for reviewing the manuscript.
This study was funded by the Armed Forces Health Surveillance Division, Global Emerging Infections Surveillance Branch, Proposal Management Information System identification nos. P0143_19_N6_01.01, P_0149_19_AH_02_NMRC and P0013_20_AH_01.01 and Navy work unit no. A1417.
G.T., D.P., J.R., C.G., J.S.A., M.S., R.Z.C., K.B.L., B.N.A., and C.D.C. are employees of the US Government. This work was prepared as part of their official duties. Title 17, U.S.C., §105 provides that copyright protection under this title is not available for any work of the US government. Title 17, U.S.C., §101 defines a US government work as a work prepared by a military service member or employee of the US government as part of that person’s official duties. The views expressed in this article reflect the results of research conducted by the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, nor the US government.
References
- Abudurexiti A, Adkins S, Alioto D, Alkhovsky SV, Avšič-Županc T, Ballinger MJ, et al. Taxonomy of the order Bunyavirales: update 2019. Arch Virol. 2019;164:1949–65. DOIPubMedGoogle Scholar
- de Carvalho MS, de Lara Pinto AZ, Pinheiro A, Rodrigues JSV, Melo FL, da Silva LA, et al. Viola phlebovirus is a novel Phlebotomus fever serogroup member identified in Lutzomyia (Lutzomyia) longipalpis from Brazilian Pantanal. Parasit Vectors. 2018;11:405. DOIPubMedGoogle Scholar
- Elliott RM, Brennan B. Emerging phleboviruses. Curr Opin Virol. 2014;5:50–7. DOIPubMedGoogle Scholar
- Gundacker ND, Carrera JP, Castillo M, Díaz Y, Valenzuela J, Tamhane A, et al. Clinical manifestations of Punta Toro virus species complex infections, Panama, 2009. Emerg Infect Dis. 2017;23:872–4. DOIPubMedGoogle Scholar
- Palacios G, Wiley MR, Travassos da Rosa APA, Guzman H, Quiroz E, Savji N, et al. Characterization of the Punta Toro species complex (genus Phlebovirus, family Bunyaviridae). J Gen Virol. 2015;96:2079–85. DOIPubMedGoogle Scholar
- Baldelli F, Ciufolini MG, Francisci D, Marchi A, Venturi G, Fiorentini C, et al. Unusual presentation of life-threatening Toscana virus meningoencephalitis. Clin Infect Dis. 2004;38:515–20. DOIPubMedGoogle Scholar
- Palacios G, Tesh R, Travassos da Rosa A, Savji N, Sze W, Jain K, et al. Characterization of the Candiru antigenic complex (Bunyaviridae: Phlebovirus), a highly diverse and reassorting group of viruses affecting humans in tropical America. J Virol. 2011;85:3811–20. DOIPubMedGoogle Scholar
- Forshey BM, Guevara C, Laguna-Torres VA, Cespedes M, Vargas J, Gianella A, et al.; NMRCD Febrile Surveillance Working Group. Arboviral etiologies of acute febrile illnesses in Western South America, 2000-2007. PLoS Negl Trop Dis. 2010;4:
e787 . DOIPubMedGoogle Scholar - Djikeng A, Halpin R, Kuzmickas R, Depasse J, Feldblyum J, Sengamalay N, et al. Viral genome sequencing by random priming methods. BMC Genomics. 2008;9:5. DOIPubMedGoogle Scholar
- Jiang H, Xing Z, Liu X, Chai Q, Xin Z, Zhu C, et al. Comparison and development of a metagenomic next-generation sequencing protocol for combined detection of DNA and RNA pathogens in cerebrospinal fluid. BMC Infect Dis. 2022;22:326. DOIPubMedGoogle Scholar
- Li PE, Lo CC, Anderson JJ, Davenport KW, Bishop-Lilly KA, Xu Y, et al. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017;45:67–80. DOIPubMedGoogle Scholar
- Zhao G, Wu G, Lim ES, Droit L, Krishnamurthy S, Barouch DH, et al. VirusSeeker, a computational pipeline for virus discovery and virome composition analysis. Virology. 2017;503:21–30. DOIPubMedGoogle Scholar
- Eifan S, Schnettler E, Dietrich I, Kohl A, Blomström AL. Non-structural proteins of arthropod-borne bunyaviruses: roles and functions. Viruses. 2013;5:2447–68. DOIPubMedGoogle Scholar
- Collao X, Palacios G, de Ory F, Sanbonmatsu S, Pérez-Ruiz M, Navarro JM, et al. Granada virus: a natural phlebovirus reassortant of the sandfly fever Naples serocomplex with low seroprevalence in humans. Am J Trop Med Hyg. 2010;83:760–5. DOIPubMedGoogle Scholar
- Amaro F, Hanke D, Zé-Zé L, Alves MJ, Becker SC, Höper D. Genetic characterization of Arrabida virus, a novel phlebovirus isolated in South Portugal. Virus Res. 2016;214:19–25. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleOriginal Publication Date: August 08, 2023
Table of Contents – Volume 29, Number 9—September 2023
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Gilda Troncos, US Naval Medical Research Unit SOUTH, 3230 Lima Pl, Washington, DC 20521-3230, USA
Top