Volume 10, Number 8—August 2004
Synopsis
Vibrio vulnificus in Taiwan
Abstract
Residents in Taiwan are often exposed to marine microorganisms through seafood and occupational exposure. The number of reported cases of infection attributable to this organism has increased since the first case was reported in 1985. The increasing number of cases may be caused by greater disease activity or improved recognition by clinicians or laboratory workers. We analyze a clinical-case series of 84 patients with V. vulnificus infection from 1995 to 2000 and describe the molecular epidemiologic features of pathogens isolated from these patients. The spectrum of clinical manifestations and outcomes, options of antimicrobial therapy, and virulence mechanisms were investigated. Results of molecular typing of isolates from humans and marine environment in this country had a high genetic divergence among these isolates. Education and measures are needed to prevent this emerging disease.
Awareness of Vibrio vulnificus as a threat to human health has evolved during the past 30 years (1). In Taiwan, Yuan et al. first reported V. vulnificus infection in a patient with septicemia and leg gangrene in Kaohsiung County in 1985 (2). Chuang et al. described an additional 27 cases during a 5-year period from May 1985 to July 1990 and demonstrated three major discernible syndromes: primary septicemia, wound infection, and gastrointestinal diseases. The disease had a high mortality rate (41%) (3). Chuang et al.’s report was also the first to demonstrate the recurrent nature of this disease. Since then, many clinicians and researchers from Taiwan have reported risk factors and the clinical spectrum of this disease on the basis of an increasing number of reported cases (4–12). Many factors have been associated with increased vulnerability of Taiwanese people to V. vulnificus infection. These include the high prevalence of hepatitis B or C virus infection-related hepatic diseases (liver cirrhosis and hepatoma), the environment, and the popularity of preparing and eating raw or undercooked seafood (3,13). These factors have drawn considerable interest to finding optimal therapeutic regimens for this infection, as well as to identifying the pathogenesis, ecology, and the reservoirs of this microorganism.
We describe the clinical features of 84 recently identified patients with V. vulnificus infection treated from 1995 to 2000 in Taiwan and report the results of molecular typing of 50 isolates of V. vulnificus from these patients. We also summarize the recent advances in understanding this newly recognized disease from the Taiwan perspective.
Figure 1

Figure 1. Estimated prevalence (per 106 population) and annual number of cases of Vibrio vulnificus infection reported from 1985 to 2000 in Taiwan. The line and triangles represent the prevalence and the bars...
Taiwan is a small island situated off the southeast coast of the Asian continent with a population of >22 million people. Figure 1 shows the annual number of reported cases and the estimated prevalence of V. vulnificus infection (per 106 persons) from 1985 to 2000 in Taiwan (2–12). Two peaks occurred: one in 1988 to 1990 (0.354–0.450/106 persons) and the other in 1996 to 2000 (0.606–1.237/106 persons). Most reported cases (>90%) occurred in residents of southern Taiwan. In Taiwan, the temperature of surface seawater is usually >18°C, except for February, when it is 17°C–23°C (6). Nearly all cases occurred in the late spring to early fall (April–October), when the seawater temperature is 20°C–29°C. The peak months for infections were June–August (summer season) when the temperature of surface seawater in Taiwan was approximately 26°C–29°C (6).
The reasons for the increased rate of V. vulnificus during the past 2 decades are not fully understood. The extent to which the increasing number of cases may be caused by increasing disease activity or improved recognition by clinicians or laboratory workers is also unclear. Since the first report of V. vulnificus infection in 1987 and subsequent reports in both humans and environmental studies, clinicians in Taiwan have become increasingly aware of the clinical features of this disease, and laboratory workers more likely to understand how to isolate and identify this pathogen accurately.
The occurrence of V. vulnificus infections in cultured shrimp and eels has been reported in Taiwan (14). A monthly survey on the distribution of Vibrionaceae in seawater from five major harbors in Taiwan was conducted from July 1991 to February 1994 (15). Among the 1,167 Vibrionaceae isolates, V. vulnificus accounted for 67 (5.7%) (15). This finding indicates that the organism exists autochthonously around the coastal waters or aquatic habitats in Taiwan. Most isolates (91%) from marine water and oysters were indole-negative (biotype I) but some belonged to biotype II (ornithine decarboxylase- and mannitol-positive) (16). Strains of V. vulnificus serovar E (also belonging to biotype II) avirulent for eels, which were recovered from water and oysters, were reported (17). Ribotyping analysis of the environmental isolates indicated a great genetic divergence among these isolates (18). More than half of the environmental isolates exhibited virulence in mice, indicating that these isolates might be pathogenic to humans (16). In addition, saline and aqueous ethanol extract (lectins) from some marine algae collected from the northeastern coast of Taiwan had marked antibacterial activity against V. vulnificus isolates recovered from the northeastern coast of Taiwan (19). Further study is needed to explore the symbiosis between marine algae and their associated marine vibrios.
Figure 2
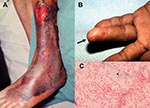
Figure 2. Characteristic skin lesions of Vibrio vulnificus infection and morphotype of the microorganism. (A) Gangrenous change with hemorrhagic bullae over the leg in a 75-year-old patient with liver cirrhosis in whom septic...
Clinical information from 84 patients V. vulnificus infection treated from 1995 to 2000 was obtained from medical records from five hospitals in Taiwan (Table). These hospitals, with a capacities of 1,500 to 2,000 beds, included National Taiwan University Hospital, Taipei; Chi-Mei Medical Center and National Cheng-Kung University Hospital, Tainan; Chang Gung Memorial Hospital-Kaohsiung, Kaohsiung; and Kaohsiung Veterans General Hospital, Kaohsiung. Most of the patients (73%) were male. More than 80% of these patients had various underlying medical conditions with liver disease (particularly hepatitis B or C virus infection-related diseases), which accounted for more than half of the patients, followed by diabetes mellitus and steroid use. Nine patients (16.3%) had exposure to marine injuries (caused by fish or crab bones or eating raw fish) or marine environments (swimming in coastal seawater or raising fish). Although 11 (20%) patients had preexisting skin wounds, exposure of the skin wounds to salt water was not known. More than 60% of these patients had a cutaneous infection, and 50% had necrotizing fasciitis. Approximately three fourths of the patients with necrotizing fasciitis had septic shock. Characteristic cutaneous lesions in patients with necrotizing fasciitis and wounds associated with bacteremia attributable to V. vulnificus are shown in Figure 2. Twenty patients (23.8%) had primary septicemia, and 3 were complicated with septic shock.
Similar to the previous findings, we found no patients with gastroenteritis caused by to V. vulnificus (3). Most patients with gastroenteritis or diarrheal illness in Taiwan do not seek care at the large teaching hospitals; they also do not usually have a stool culture, which might explain the lack of patients with gastrointestinal illness attributable to V. vulnificus.
Figure 3
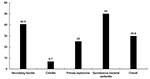
Figure 3. Rates of deaths according to different types of infection of 84 patients with Vibrio vulnificus infection.
A third-generation cephalosporin plus minocycline was used as the definite treatment regimen in 46% of patients. Among 57 patients with cutaneous lesions, 49 (86.0%) had some form of surgical treatment (incision and drainage, débridement, fasciotomy, and amputation). The overall case-fatality rate was approximately 30% (Figure 3), which was similar to that reported previously among patients seen from 1995 to 1990 (3). Patients with spontaneous bacterial peritonitis had the highest case-fatality rate (50%), followed by necrotizing fasciitis (40.5%). Patients with cellulitis had the lowest case-fatality rate (6.7%).
MICs were determined and interpreted by using the MIC interpretive criteria for Enterobacteriaceae recommended by the National Committee for Clinical Laboratory Standards (20–23). All isolates of V. vulnificus in Taiwan, which were collected from the previous studies, were susceptible to the following agents (MIC90): ampicillin (1 μg/mL), carbenicillin (4 μg/mL), cephalothin (4 μg/mL), cefamandole (2 μg/mL), cefotaxime (<0.03–0.06 μg/mL), ceftriaxone (<0.03 μg/mL), cefoperazone (0.12 μg/mL), aztreonam (8 μg/mL), imipenem (<0.03–0.12 μg/mL), gentamicin (4 μg/mL), amikacin (8 μg/mL), tetracycline (0.25 μg/mL), minocycline (0.06–0.25 μg/mL), chloramphenicol (0.5 μg/mL), and fluoroquinolones: ofloxacin (<0.03 μg/mL), lomefloxacin (0.12 μg/mL), ciprofloxacin (<0.03-0.03 μg/mL), levofloxacin (0.03 μg/mL), moxifloxacin (0.06 μg/mL), gatifloxacin (0.06 μg/mL), and sparfloxacin (0.06 μg/mL) (20–23). Few isolates were not susceptible to ceftazidime (MIC 32 μg/mL) and moxalactam (MIC 32 μg/mL) (21). All isolates were resistant to clindamycin (MICs >256 μg/mL) (20). In vitro synergism between cefotaxime and minocycline against V. vulnificus isolates was documented by time-kill study (21). Time-kill study also demonstrated that fluoroquinolones at concentrations of two times the MIC had a persistent inhibitory effect on V. vulnificus for >48 hours (23).
In vivo study using a mouse model of V. vulnificus infection clearly indicated that combination therapy with cefotaxime and minocycline is distinctly superior to therapy with cefotaxime or minocycline alone (22). A similar effect of newer fluoroquinolones as single agents compared with the cefotaxime-minocycline combination was also demonstrated in the treatment of severe experimental V. vulnificus infection (23).
On the basis of the in vitro and in vivo animal studies, along with clinical outcome analysis, combination therapy with cefotaxime (2 g every 6 h intravenously) and minocycline (100 mg every 12 h intravenously) was recommended for treating adult patients with bacteremia and severe soft-tissue infection caused by V. vulnificus (21,22). For severe soft-tissue infection (necrotizing fasciitis, tissue necrosis with gangrene change, and myositis), early and aggressive surgical interventions (incision and drainage, débridement, fasciotomy, and amputation) are important in saving the life of the patient.
More than 90% of V. vulnificus isolates whose biotypes were determined belonged to biotype I, which is well known to be pathogenic for humans (15,16). In 1997, Chuang et al. first demonstrated that severe damage of the connective tissue of a mouse by V. vulnificus wound infection could be mediated by a recombinant extracellular metalloprotease (able to digest collagen and elastin) (24). Lee et al. also illustrated that extracellular products of V. vulnificus were lethal to fish (moribund black porgy, Acanthopagrus schlegeli) (25). Genes (vvp and empV) encoding the metalloprotease and gene (vllY) encoding a novel hemolysin of V. vulnificus were subsequently cloned and characterized (26–28).
Hor et al. showed that isogenic protease-deficient (PD) mutant of V. vulnificus was as virulent as its parent strains in mice infected intraperitoneally and was 10-fold more virulent in mice infected through the oral route (29). A metalloprotease- and cytolysin-deficient mutant of V. vulnificus also had similar virulence in mice, and its cytotoxicity for HEP-2 cells (cytotoxin) compared with those of the wild-type strains (30). These findings suggest that neither metalloprotease nor cytolysin is essential for the virulence or invasiveness of V. vulnificus in mice. A possible multifactor interaction in bacterial virulence might be present but to an extent that is not yet clear. However, two genes, vvn (encoding a periplasmic nuclease, Vvn) and smcR (encoding SmcR, which regulat metalloprotease gene expression), were not required for V. vulnificus virulence in mice (31,32).
Animal studies clearly demonstrated that iron could increase the growth rate of V. vulnificus, which quickly reached a lethal concentration with enhanced cytotoxicity in the iron-overloaded mice (33). A study of the survival of V. vulnificus in whole blood from patients with different degrees of liver disease showed that high serum ferritin levels and low phagocytosis activity of neutrophils were independent and important predictors of survival of the organism in blood (34). These findings indicated that patients with chronic hepatitis, liver cirrhosis, and hepatoma (high serum ferritin levels and lower phagocytosis) were at high risk for V. vulnificus infection (34). Although many putative virulence factors have been studied for this exceptionally virulent human pathogen in Taiwan, how these factors and other veiled factors (such as capsular polysaccharide and lipopolysaccharide) interact to produce dramatic infections and what host aspects (such as overproduction of proinflammatory cytokines) are essential to infection are yet to be elucidated (3).
Results of molecular typing by using restriction fragment length polymorphism analysis of rRNA (ribotyping) among 13 clinical and environmental (from seawater and eels in southern Taiwan) isolates of V. vulnificus and arbitrarily primed polymerase chain reaction analysis of 37 isolates (24 clinical isolates and 13 from seawater from coast areas around Taiwan) were previously reported (18,35). Both showed high genetic divergence among clinical and environmental isolates.
The concentration of V. vulnificus in recent clinical and environmental isolates in southern Taiwan indicates the possibility of clonal spread in this area. In this study, 50 isolates of V. vulnificus collected from 1995 to 2000 from southern (46 isolates) and northern (4 isolates) Taiwan were analyzed. These isolates included those from various clinical specimens (blood and wound pus) of 50 patients with V. vulnificus infection. All isolates of V. vulnificus were identified by using conventional methods and the O/129 susceptibility tests. Identification of the isolates was further confirmed by the API 32 GN system (bioMérieux Vitek, Inc., Hazelwood, MO). Pulsed-field gel electrophoresis (PFGE) analysis was performed by a method described previously by Tenover et al. (36,37). DNA was digested by the restriction enzymes SfiI and NotI (Promega, Madison, WI). All isolates were not identical in PFGE profiles (50 pulsotypes were found), and only two isolates from southern Taiwan were closely related (within three bands of difference). These findings support the high degree of heterogeneity among isolates of V. vulnificus that cause human infections in Taiwan.
Residents of Taiwan, particularly those with preexisting liver and other chronic, underlying medical conditions (renal disease, diabetes mellitus, chronic steroid use), should be educated in measures to prevent acquiring V. vulnificus infections. This bacterium is present in warm coastal waters around Taiwan during the summer months, particularly in the southern region. Exposing open wounds or broken skin to warm salt or brackish water or to raw marine animals harvested from such waters should be avoided. Patients at high risk should wear protective clothing (e.g., gloves) when handling seafood (fish, oysters, clams, shrimp, eels, and other shellfish) and not eat raw or improperly cooked seafood. Because this disease is rapidly progressive and deadly if not recognized promptly and treated aggressively, any illness (such as fever or skin lesions), which develops in patients at risk after contact with marine animals or waters or ingestion of seafood requires immediate medical care.
The government in Taiwan (Departments of Health and Council of Agriculture) should encourage food companies to put warning labels on seafood containers, menus, and public health brochures. The wording of such labeling should be similar to the label required by the Florida Department of Natural Resources for all wholesale shell food and shucked products: “Consumer Information—There is a risk associated with consuming raw oysters or any raw animal protein. If you have chronic illness of the liver, stomach, or blood or have immune disorders, you are at a greater risk of serious illness from raw oysters and should eat oysters fully cooked. If unsure of your risk, consult a physician” (38).
Residents of Taiwan have a high prevalence of chronic liver disease and are often exposed to marine microorganisms present in the sea that surrounds the island or rivers, lakes, or ponds inside the island. The presence of high genetic divergence among V. vulnificus isolates from humans and the environment indicates that this virulent bacterium is ubiquitous in nature. When V. vulnificus is suspected as the cause of sepsis, empiric therapy that includes a third-generation cephalosporin and minocycline should be administered. It should be standard practice for physicians to advise patients with underlying medical illness to avoid eating raw or undercooked seafood and to avoid exposing wounds to seawater.
Dr. Hsueh is an associate professor in the departments of Laboratory Medicine and Internal Medicine of National Taiwan University Hospital and National Taiwan University College of Medicine. His research interests include mechanisms of antimicrobial resistance and molecular epidemiology of emerging pathogens.
References
- Roland FP. Leg gangrene and endotoxin shock due to vibrio parahaemolyticus—an infection acquired in New England coastal waters. N Engl J Med. 1970;282:1306. DOIPubMedGoogle Scholar
- Yuan CY, Yuan CC, Wei DC, Lee AM. Septicemia and gangrenous change of the legs caused by marine vibrio, Vibrio vulnificus—report of a case. Taiwan Yi Xue Hui Za Zhi. 1987;86:448–51.PubMedGoogle Scholar
- Chuang YC, Yuan CY, Liu CY, Lan CK, Huang AH. Vibrio vulnificus infection in Taiwan: report of 28 cases and review of clinical manifestations and treatment. Clin Infect Dis. 1992;15:271–6.PubMedGoogle Scholar
- Chuang YC, Young CD, Chen CW. Vibrio vulnificus infection. Scand J Infect Dis. 1989;21:721–6. DOIPubMedGoogle Scholar
- Lee SY, Chuang YC, Young CD. Extensive cellulitis with septic shock caused by Vibrio vulnificus infection—a case report with review of literature. Kansenshogaku Zasshi. 1991;65:1484–7.PubMedGoogle Scholar
- Chao CH, Duh RW, Liu CY, Lou JP, Chen CK. Experience of six patients with Vibrio vulnificus septicemia. Zhonghua Yi Xue Za Zhi (Taipei). 1992;49:335–42.PubMedGoogle Scholar
- Chang JJ, Sheen IS, Peng SM, Chen PC, Wu CS, Leu HS. Vibrio vulnificus infection—report of 8 cases and review of cases in Taiwan. Changgeng Yi Xue Za Zhi. 1994;17:339–46.PubMedGoogle Scholar
- Shih YT, Peng CT, Tsai CH, Tsai FJ. β-thalassemia major complicated with Vibrio vulnificus septicemia. Zhonghua Min Guo Xiao Er Ke Yi Xue Hui Za Zhi. 1994;35:84–9.PubMedGoogle Scholar
- Tsai WC, Liu YC, Yen MY, Wang JH, Chen YS, Wang JH, Vibrio vulnificus infections: experience of thirteen cases in southern Taiwan. J Microbiol Immunol Infect. 1998;31:46–50.
- Wang SM, Liu CC, Chiou YY, Yang HB, Chen CT. Vibrio vulnificus infection complicated by acute respiratory distress syndrome in a child with nephrotic syndrome. Pediatr Pulmonol. 2000;29:400–3. DOIPubMedGoogle Scholar
- Wu CH, Hu WH, Hung DZ, Peng YC, Yang DY. Snakebite complicated with Vibrio vulnificus infection. Vet Hum Toxicol. 2001;43:283–5.PubMedGoogle Scholar
- Chiu S, Chiu CH, Jaing TH, Chang KJ, Lin TY. Necrotising fasciitis caused by Vibrio vulnificus in a child without known risk factors. Eur J Pediatr. 2002;161:464–5. DOIPubMedGoogle Scholar
- Chiang SR, Chuang YC. Vibrio vulnificus infection: clinical manifestations, pathogenesis, and antimicrobial therapy. J Microbiol Immunol Infect. 2003;36:81–8.PubMedGoogle Scholar
- Song YL, Cheng W, Shen CH, Ou YC, Song HB. Occurrence of Vibrio vulnificus infection in cultured shrimp and eel in Taiwan. National Science Council Symposium Series. 1990;16:172–9.
- Wu HS, Liu DP, Hwang CH, Chen MJ, Hwang JL, Liu Y, Survey on the distribution of Vibrionaceae at the seaport areas in Taiwan, 1991–1994. Zhonghua Min Guo Wei Sheng Wu Ji Mian Yi Xue Za Zhi. 1996;29:197–209.PubMedGoogle Scholar
- Hor LI, Gao CT, Wan L. Isolation and characterization of Vibrio vulnificus inhabiting the marine environment of the southwestern area of Taiwan. J Biomed Sci. 1995;2:384–9. DOIPubMedGoogle Scholar
- Amaro C, Hor LI, Marco-Noales E, Bosque T, Fouz B, Alcaide E. Isolation of Vibrio vulnificus serovar E from aquatic habitats in Taiwan. Appl Environ Microbiol. 1999;65:1352–5.PubMedGoogle Scholar
- Yang SF, Cheng JC, Hor LI. Ribotyping of clinical Vibrio vulnificus isolates. Zhonghua Min Guo Wei Sheng Wu Ji Mian Yi Xue Za Zhi. 1995;28:270–9.PubMedGoogle Scholar
- Liao WR, Lin JY, Shieh WY, Jeng WL, Huang R. Antibiotic activity of lectins from marine algae against marine vibrios. J Ind Microbiol Biotechnol. 2003;30:433–9. DOIPubMedGoogle Scholar
- Hsueh PR, Chang JC, Chang SC, Ho SW, Hsieh WC. In vitro antimicrobial susceptibility of Vibrio vulnificus isolated in Taiwan. Eur J Clin Microbiol Infect Dis. 1995;14:151–3. DOIPubMedGoogle Scholar
- Chuang YC, Liu JW, Ko WC, Lin KY, Wu JJ, Huang KY. In vitro synergism between cefotaxime and minocycline against Vibrio vulnificus. Antimicrob Agents Chemother. 1997;41:2214–7.PubMedGoogle Scholar
- Chuang YC, Ko WC, Wang ST, Liu JW, Kuo CF, Wu JJ, Minocycline and cefotaxime in the treatment of experimental murine Vibrio vulnificus infection. Antimicrob Agents Chemother. 1998;42:1319–22.PubMedGoogle Scholar
- Tang HJ, Chang MC, Ko WC, Huang KY, Lee CY, Chuang YC. In vitro and in vivo activities of newer fluoroquinolones against Vibrio vulnificus. Antimicrob Agents Chemother. 2002;46:3580–4. DOIPubMedGoogle Scholar
- Chuang YC, Sheu HM, Ko WC, Chang TM, Chang MC, Huang KY. Mouse skin damage caused by a recombinant extracellular metalloprotease from Vibrio vulnificus and by V. vulnificus infection. J Formos Med Assoc. 1997;96:677–84.PubMedGoogle Scholar
- Lee KK, Chiang HT, Yii KC, Su WM, Liu PC. Effects of extracellular products of Vibrio vulnificus on Acanthopagrus schlegeri serum components in vitro and in vivo. Microbios. 1997;92:209–17.PubMedGoogle Scholar
- Cheng JC, Shao CP, Hor LI. Cloning and nucleotide sequencing of the protease gene of Vibrio vulnificus. Gene. 1996;183:255–7. DOIPubMedGoogle Scholar
- Chuang YC, Chang TM, Chang MC. Cloning and characterization of the gene (empV) encoding extracellular metalloprotease from Vibrio vulnificus. Gene. 1997;189:163–8. DOIPubMedGoogle Scholar
- Chang TM, Chuang YC, Su JH, Chang MC. Cloning and sequence analysis of a novel hemolysin gene (vllY) from Vibrio vulnificus. Appl Environ Microbiol. 1997;63:3851–7.PubMedGoogle Scholar
- Shao CP, Hor LI. Metalloprotease is not essential for Vibrio vulnificus virulence in mice. Infect Immun. 2000;68:3569–73. DOIPubMedGoogle Scholar
- Fan JJ, Shao CP, Ho YC, Yu CK, Hor LI. Isolation and characterization of a Vibrio vulnificus mutant deficient in both extracellular metalloprotease and cytolysin. Infect Immun. 2001;69:5943–8. DOIPubMedGoogle Scholar
- Shao CP, Hor LI. Regulation of metalloprotease gene expression in Vibrio vulnificus by a Vibrio harveyi LuxR homologue. J Bacteriol. 2001;183:1369–75. DOIPubMedGoogle Scholar
- Wu SI, Lo SK, Shao CP, Tsai HW, Hor LI. Cloning and characterization of a periplasmic nuclease of Vibrio vulnificus and its role in preventing uptake of foreign DNA. Appl Environ Microbiol. 2001;67:82–8. DOIPubMedGoogle Scholar
- Hor LI, Chang YK, Chang CC, Lei HY, Ou JT. Mechanism of high susceptibility of iron-oveloaded mouse to Vibrio vulnificus infection. Microbiol Immunol. 2000;44:871–8.PubMedGoogle Scholar
- Hor LI, Chang TT, Wang ST. Survival of Vibrio vulnificus in whole blood from patients with chronic liver diseases: association with phagocytosis by neutrophils and serum ferritin levels. J Infect Dis. 1999;179:275–8. DOIPubMedGoogle Scholar
- Wu JJ, Hor LI, Shiau SL. Differentiation of Vibrio vulnificus strains by an arbitrarily primed polymerase chain reaction. Zhonghua Min Guo Wei Sheng Wu Ji Mian Yi Xue Za Zhi. 1995;28:70–8.PubMedGoogle Scholar
- Tamplin ML, Jackson JK, Buchrieser C, Murphree RL, Portier KM, Gangar V, Pulsed-field gel electrophoresis and ribotype profiles of clinical and environmental Vibrio vulnificus isolates. Appl Environ Microbiol. 1996;62:3572–80.PubMedGoogle Scholar
- Tenover FC, Arbeit R, Goering RV, Mickelsen PA, Murray BE, Persing DH, Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–9.PubMedGoogle Scholar
- Centers for Diseases Control and Prevention. Vibrio vulnificus infections associated with raw oyster consumption—Florida, 1981–1992. MMWR Morb Mortal Wkly Rep. 1993;42:405–7.PubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 10, Number 8—August 2004
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Po-Ren Hsueh, National Taiwan University Hospital, No. 7, Chung-Shan South Road, Taipei; fax: 886-2-232-4263; ; and Yin-Ching Chuang, Chi-Mei Medical Center, 901 Chung-Hwa Road, Yung-kang City, Tainan, Taiwan 710; fax: 886-6-251-7849Po-Ren Hsueh, National Taiwan University Hospital, No. 7, Chung-Shan South Road, Taipei; fax: 886-2-232-4263; ; and Yin-Ching Chuang, Chi-Mei Medical Center, 901 Chung-Hwa Road, Yung-kang City, Tainan, Taiwan 710; fax: 886-6-251-7849
Top