Volume 14, Number 1—January 2008
Dispatch
Prolonged Bartonella henselae Bacteremia Caused by Reinfection in Cats
Abstract
We analyzed the genetic relatedness of blood culture isolates of Bartonella henselae from 2 cats of patients with cat-scratch disease at admission and after 12 months. Isolates from each cat at different times were clonally unrelated, which suggested reinfection by a second strain.
Bartonella henselae is a zoonotic pathogen associated with a broad spectrum of disease manifestations in humans. Cat-scratch disease (CSD) is commonly encountered in immunocompetent patients; in immunocompromised patients, bacillary angiomatosis, peliosis hepatis, and recurrent bacteremia are usually seen. Domestic cats represent the main host and reservoir for B. henselae (1). Recurrent, intraerythrocytic bacteremia develops in infected cats without overt clinical symptoms (2). Experimental infection of specific pathogen–free cats with B. henselae induces recurrent episodes of bacteremia, which, in most cases, resolve spontaneously within 22–33 weeks postinfection (3–5). Prolonged bacteremia >7 months has been documented sporadically, e.g., in 1 of 12 experimentally infected cats inoculated with the highest infectious dose (this cat was bacteremic 32 weeks postinfection) (3), or in an unspecified number of cases in 21 experimentally infected cats that were bacteremic 48 weeks postinfection (5).
Few studies have investigated the course of recurrent bacteremia in naturally infected cats because follow-up investigations are difficult to conduct (1,6–9). Koehler et al. (1) detected recurrent bacteremia with a duration >2 months in 3 cats of patients with bacillary angiomatosis. Kordick et al. (6) reported positive blood cultures in cats of several CSD patients up to 14 months after collection of the initial positive culture. In the latter study, the first blood culture was collected from the index cat of 1 CSD patient 22 months after the onset of the disease in the patient and contained B. henselae (7). Sander et al. (8) found repeated bacteremia in the cat of a CSD patient after 5 months and in 2 other cats after 1 year. In another study, B. henselae was isolated from the blood culture of a cat of a patient who had an episode 2.5 years earlier of debilitating fatigue with a duration of 1 month and without fever or lymphadenopathy (7). B. henselae was isolated again from the blood culture of the index cat after 5 months (7).
In these studies, the question whether the cats were still infected by the initial B. henselae strain or had acquired a new strain was not addressed. It was concluded that the cats were persistently infected with B. henselae (8). We have recently demonstrated the appropriateness of pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) for differentiation of B. henselae isolates to the strain level (10,11). Therefore, we analyzed the clonal relationship between sequential B. henselae isolates that were obtained at different times from the blood of 2 cats to determine whether recurrences were caused by the initial strain or a new strain.
Figure
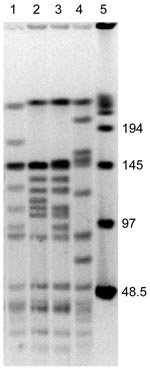
Figure. SmaI macrorestriction patterns of Bartonella henselae isolates from 2 cats. Lane 1, cat 36, first isolate; lane 2, cat 36, second isolate obtained 12 months later; lane 3, cat 75, first...
We tested 4 isolates of B. henselae: FR96/BK36, FR96/BK36II, FR96/BK75, and FR96/BK75II. These isolates were grown from the blood of 2 naturally infected cats (cat 36 and cat 75) of CSD patients at first consultation and after 12 months, respectively (9). The original colony counts were 100, 100, 120, and 100, respectively. PFGE analysis was conducted after digestion of DNA with SmaI, and MLST was conducted after partial sequencing of 8 genetic loci (12,13). PFGE analysis showed 9 band differences between isolates 36 and 36II and 10 band differences between isolates 75 and 75II (Figure), which suggested that isolates obtained from the same cat at different times were not clonally related (14). MLST analysis showed 3 and 6 different alleles between isolates 36 and 36II and isolates 75 and 75II, respectively (Table). Isolates 36 and 36II were assigned to sequence type (ST) 14 and ST5, which have the 16S rRNA alleles 1 and 2, respectively. Isolates 75 and 75II were assigned to ST5 and ST7, respectively, and both had the 16S rRNA allele 2 (Table). These data suggest that both cats were infected by a second B. henselae strain at the second time blood was obtained.
Our data indicate that recurrent B. henselae bacteremia in naturally infected cats does not necessarily represent a relapse but may be caused by reinfection. We were surprised to find that both cats were presumably reinfected by a different strain within 1 year. The interval between collection of the initial and follow-up blood samples was long, which might explain the high rate of reinfection. The possibility that the cats were infected by 2 B. henselae stains at the time blood was first obtained was examined by subjecting 5 single-colony–derived cultures of each initial isolate to PFGE analysis, which did not show evidence for co-infection. In addition, co-infection by 2 strains is unlikely because we would not have been able to determine a unique sequence for those genetic loci that displayed allelic polymorphism. Furthermore, co-infection by 2 strains in both cats would represent a rare coincidence. Nevertheless, we cannot completely rule out co-infection with 2 strains at the time blood was first obtained, with 1 strain being highly underrepresented among initial colonies.
To our knowledge, this is the first report to document sequential infection by different B. henselae strains in naturally infected cats by using 2 independent typing methods. In a previous study, Kabeya et al. (9) reported variations of up to 5 bands in PFGE patterns of B. henselae isolates obtained from naturally infected cats during different bacteremic peaks. However, we have recently demonstrated that genetic variants displaying 1- to 4-band differences frequently occur within primary B. henselae isolates and do not necessarily indicate infection by a different strain (11,12). Careful interpretation of PFGE typing results, use of additional restriction endonucleases, or use of other typing techniques is necessary to ensure correct classification of different patterns to the variant or strain level. B. henselae isolates collected from different bacteremic episodes of naturally infected cats after a long interval should by tested by molecular typing to determine their clonal relatedness.
In conclusion, our data emphasize the requirement for molecular typing to differentiate between relapse and reinfection by B. henselae in naturally infected cats. Studies on additional isolates are required to evaluate the frequency of reinfection by a different strain in naturally infected cats. Results of these studies would provide a better understanding of the natural course of feline infection. Our data also suggest that infection by a distinct B. henselae strain does not induce protective immunity against subsequent infection by a clonally unrelated strain. These results are partially consistent with those of Yamamoto et al. (15), who found incomplete cross-protection between isolates with different 16S rRNA alleles. Recent studies have shown that the delineation of B. henselae isolates into 2 genotypes on the basis of 16S rRNA sequence is not consistent with phylogenetic classifications using other genetic loci and does not reflect clonal lineage of isolates (11,14). Reevaluation of induction of cross-protection between different B. henselae strains in an experimental infection model is needed.
Dr Arvand is a professor of medical microbiology at the Institute for Medical Microbiology, Virology and Hygiene, University of Rostock, Rostock, Germany. Her research interests include pathogenesis, persistence, and clinical and epidemiologic aspects of B. henselae infections.
Acknowledgments
We thank Anna Sander for providing B. henselae isolates.
This study was supported by a grant from the Deutsche Forschungsgemeinschaft to M.A.
References
- Koehler JE, Glaser CA, Tappero JW. Rochalimaea henselae infection. A new zoonosis with the domestic cat as reservoir. JAMA. 1994;271:531–5. DOIPubMedGoogle Scholar
- Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. 1996;34:1952–6.PubMedGoogle Scholar
- Guptill L, Slater L, Wu CC, Lin TL, Glickman LT, Welch DF, Experimental infection of young specific pathogen-free cats with Bartonella henselae. J Infect Dis. 1997;176:206–16.PubMedGoogle Scholar
- O’Reilly KL, Bauer RW, Freeland RL, Foil LD, Hughes KJ, Rohde KR, Acute clinical disease in cats following infection with a pathogenic strain of Bartonella henselae (LSU16). Infect Immun. 1999;67:3066–72.PubMedGoogle Scholar
- Yamamoto K, Chomel BB, Kasten RW, Hew CM, Weber DK, Lee WI. Experimental infection of specific pathogen free (SPF) cats with two different strains of Bartonella henselae type I: a comparative study. Vet Res. 2002;33:669–84. DOIPubMedGoogle Scholar
- Kordick DL, Wilson KH, Sexton DJ, Hadfield TL, Berkhoff HA, Breitschwerdt EB. Prolonged Bartonella bacteremia in cats associated with cat-scratch disease patients. J Clin Microbiol. 1995;33:3245–51.PubMedGoogle Scholar
- Kordick DL, Breitschwerdt EB. Persistent infection of pets within a household with three Bartonella species. Emerg Infect Dis. 1998;4:325–8.PubMedGoogle Scholar
- Sander A, Buhler C, Pelz K, von Cramm E, Bredt W. Detection and identification of two Bartonella henselae variants in domestic cats in Germany. J Clin Microbiol. 1997;35:584–7.PubMedGoogle Scholar
- Kabeya H, Maruyama S, Irei M, Takahashi R, Yamashita M, Mikami T. Genomic variations among Bartonella henselae isolates derived from naturally infected cats. Vet Microbiol. 2002;89:211–21. DOIPubMedGoogle Scholar
- Arvand M, Viezens J. Evaluation of pulsed-field gel electrophoresis and multi-locus sequence typing for the analysis of clonal relatedness among Bartonella henselae isolates. Int J Med Microbiol. 2007;297:255–62. Epub 2007 Mar 30. DOIPubMedGoogle Scholar
- Berghoff J, Viezens J, Guptill L, Fabbi M, Arvand M. Bartonella henselae exists as a mosaic of different genetic variants in the infected host. Microbiology. 2007;153:2045–51. DOIPubMedGoogle Scholar
- Arvand M, Schubert H, Viezens J. Emergence of distinct genetic variants in the population of primary Bartonella henselae isolates. Microbes Infect. 2006;8:1315–20. DOIPubMedGoogle Scholar
- Iredell J, Blanckenberg D, Arvand M, Grauling S, Feil EJ, Birtles RJ. Characterization of the natural population of Bartonella henselae by multilocus sequence typing. J Clin Microbiol. 2003;41:5071–9. DOIPubMedGoogle Scholar
- Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–9.PubMedGoogle Scholar
- Yamamoto K, Chomel BB, Kasten RW, Hew CM, Weber DK, Lee WI, Infection and re-infection of domestic cats with various Bartonella species or types: B. henselae type I is protective against heterologous challenge with B. henselae type II. Vet Microbiol. 2003;92:73–86. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleTable of Contents – Volume 14, Number 1—January 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Mardjan Arvand, Institut für Medizinische Mikrobiologie, Virologie und Hygiene, Universität Rostock, Schillingallee 70, D-18057 Rostock, Germany;
Top