Volume 15, Number 10—October 2009
CME ACTIVITY - Research
Community-Associated Methicillin-Resistant Staphylococcus aureus, Iowa, USA
Introduction
![]()
MedscapeCME is pleased to provide online continuing medical education (CME) for this journal article, allowing clinicians the opportunity to earn CME credit. This activity has been planned and implemented in accordance with the Essential Areas and policies of the Accreditation Council for Continuing Medical Education through the joint sponsorship of MedscapeCME and Emerging Infectious Diseases. MedscapeCME is accredited by the Accreditation Council for Continuing Medical Education (ACCME) to provide continuing medical education for physicians. MedscapeCME designates this educational activity for a maximum of 0.5 AMA PRA Category 1 Credits™. Physicians should only claim credit commensurate with the extent of their participation in the activity. All other clinicians completing this activity will be issued a certificate of participation. To participate in this journal CME activity: (1) review the learning objectives and author disclosures; (2) study the education content; (3) take the post-test and/or complete the evaluation at http://www.medscape.com/cme/eid; (4) view/print certificate.
Learning Objectives
• Upon completion of this activity, participants will be able to:
• Describe risk factors for healthcare-associated methicillin-resistant Staphylococcus aureus (MRSA) infection
• Define multiresistant MRSA isolates
• Describe characteristics of patients who are likely to be infected by strains USA300/400 of MRSA
• Identify predictive factors for community-associated MRSA infection in 1 US state
• Describe recent patterns of community-associated MRSA infection in the United States
Editor
Beverly Merritt, Technical Writer-Editor, Emerging Infectious Diseases. Disclosure: Beverly Merritt has disclosed no relevant financial relationships.
CME AUTHOR
Désirée Lie, MD, MSEd, Clinical Professor, Family Medicine, University of California, Orange; Director, Division of Faculty Development, UCI Medical Center, Orange, California. Disclosure: Désirée Lie, MD, MSEd, has disclosed no relevant financial relationships.
AUTHORS
Philip Van De Griend, MPH; Loreen A. Herwaldt, MD; Bret Alvis, MD; Mary DeMartino, BS, MT, (ASCP)SM; Kristopher Heilmann, BS; and Diana DeSalvo Vonstein, BS, MPH, have disclosed no relevant financial relationships. Gary Doern, MD, has disclosed that he has served as an advisor or consultant to Astellas Pharma, Inc. and has served as a speaker or a member of a speaker's bureau for Pfizer Inc.; Cubist Pharmaceuticals; Schering-Plough Corporation; and Astellas Pharma, Inc. Patricia Winokur, MD, has disclosed that she has received grants for clinical research from CSL Limited and owns stock, stock options, or bonds from Johnson & Johnson Pharmaceutical Research & Development L.L.C. Daniel Diekema, MD, MS, has disclosed that he has received grants for clinical research from Merck & Co., Inc.; Pfizer Inc.; Schering-Plough Corporation; and Astellas Pharma, Inc.
Abstract
We performed antimicrobial drug susceptibility testing and molecular typing on invasive methicillin-resistant Staphylococcus aureus (MRSA) isolates (n = 1,666) submitted to the University of Iowa Hygienic Laboratory during 1999–2006 as part of a statewide surveillance system. All USA300 and USA400 isolates were resistant to <3 non–β-lactam antimicrobial drug classes. The proportion of MRSA isolates from invasive infections that were either USA300 or USA400 increased significantly from 1999–2005 through 2006 (p<0.0001). During 2006, the incidence of invasive community-associated (CA)–MRSA infections was highest in the summer (p = 0.0004). Age <69 years was associated with an increased risk for invasive CA-MRSA infection (odds ratio [OR] 5.1, 95% confidence interval [CI] 2.06–12.64), and hospital exposure was associated with decreased risk (OR 0.07, 95% CI 0.01–0.51).
Methicillin-resistant Staphylococcus aureus (MRSA) emerged in the 1960s and has since become a major cause of illness and death in the healthcare setting (1,2). Risk factors for infection with healthcare-associated MRSA (HA-MRSA) include hospitalization, residence in a long-term care facility, older age, invasive devices (e.g., catheters, feeding tubes), and exposure to antimicrobial agents. HA-MRSA isolates are often resistant to several antimicrobial drug classes in addition to β-lactams (3).
In the 1990s, investigators began describing serious MRSA infections among persons who did not have typical risk factors for infections with this organism (2,4–8). These community-associated MRSA (CA-MRSA) infections affected young, healthy persons (4,5,7) and were associated with factors such as participating in contact sports, sharing towels or athletic equipment, using illegal intravenous drugs, and living in crowded or unsanitary areas (e.g., prisons, hurricane evacuee centers) (9,10).
Pulsed-field gel electrophoresis (PFGE) demonstrated that MRSA strains causing these community-associated infections (USA300 and USA400) were different than those causing healthcare-associated infections (USA100 and USA200) (11). USA300 and USA400 MRSA strains typically have the staphylococcal cassette chromosome (SCC) mec type IV, not the SCCmec type II carried by most USA100 and USA200 isolates (12). In addition, USA300/400 isolates usually carry the gene that encodes the Panton-Valentine leukocidin (pvl), a bicomponent (lukF-PV and lukS-PV) pore-forming leukotoxin (8,13–15). Currently, the role of PVL in the pathogenesis of infections caused by USA300/400 isolates is controversial. Epidemiologic studies and a study by Labandeira-Rey et al. suggest that PVL is associated with virulence and causes the necrosis characteristic of infections with these strains (16). In contrast, a study by Voyich et al. found no difference in virulence between the wild-type parent strains and the isogenic knockout strains that did not produce PVL (17).
A recent multicenter study by Moran et al. showed that USA300 MRSA is now the most common cause of skin and soft tissue infections (SSTIs) among adults seeking treatment in emergency departments in 11 large metropolitan areas (15). USA300 also causes serious invasive infections such as necrotizing pneumonia, bloodstream infections, and surgical site infections, some of which are acquired in hospitals (18–22). Although most USA300 and USA400 isolates are currently resistant to fewer classes of antimicrobial drugs than are HA-MRSA isolates (13), a recent paper by Han et al. identified a USA300 subtype that is resistant to erythromycin, clindamycin (constitutive), tetracycline, mupirocin, and fluoroquinolones (23).
Most epidemiologic studies of CA-MRSA have examined isolates from SSTIs infections (7,15,18), and most studies that evaluated patients with invasive disease have involved single healthcare facilities (21,24) or isolates obtained primarily from large urban areas (22). We describe the molecular epidemiology of invasive infections caused by USA300 and USA400 in a rural state. We characterized invasive MRSA from 1999–2005 (select isolates) and in 2006 (all isolates) submitted to the statewide surveillance system in Iowa for invasive MRSA infections.
As part of a statewide surveillance system, the Iowa Department of Public Health has mandated since 1999 that clinical microbiology laboratories submit invasive isolates of MRSA to the University Hygienic Laboratory (UHL), Iowa’s public health laboratory (25,26). After performing antimicrobial drug susceptibility testing on all isolates, we further characterized (by PFGE, PVL detection, and SCCmec typing) all isolates from 1999–2005 that were resistant to <3 non–β-lactam antimicrobial drug classes (i.e., consistent with USA300/400) and all 343 isolates from unique patients with invasive infections submitted to the UHL during 2006.
Antimicrobial Drug Susceptibility Testing
All invasive MRSA isolates during 1999–2006 were tested for antimicrobial drug susceptibility by the broth dilution method described by the Clinical and Laboratory Standards Institute (27). Invasive isolates were defined as any organism from a normally sterile body site such as blood, cerebrospinal fluid, pleural fluid, joint fluid, or fluid from a liver abscess. Isolates from urine were excluded.
Isolates were tested for susceptibility to tetracycline, erythromycin, clindamycin, trimethoprim/sulfamethoxazole, gentamicin, levofloxacin, moxifloxacin, linezolid, daptomycin, vancomycin, and rifampin. Multidrug-resistant isolates were defined as MRSA isolates that were resistant to more than 3 of 8 representative antimicrobial drug classes: macrolides (erythromycin), lincosamides (clindamycin), quinolones (levofloxacin or moxifloxacin), tetracyclines, sulfa drugs (trimethoprim/sulfamethoxazole), aminoglycosides (gentamicin), glycopeptides (vancomycin), and rifampin.
Molecular Typing and PCR to Assess SCCmec Type and Presence of the PVL Gene
PFGE was performed as previously described (28). Each gel accommodated bacteriophage Lambda ladders (at 3 places on the gel), DNA from 17 isolates, type strains for USA300 and USA400 from the Centers for Disease Control and Prevention (Atlanta, GA, USA), and S. aureus NCTC-8325 (at 3 places on the gel). The gel images were saved as TIFF files and BioNumerics computer software (Biosystematica, Llandysul, Wales, UK) was used to perform cluster analysis. Isolates were classified as the same strain if cluster analysis indicated that they were >80% similar. PFGE patterns for clinical isolates were compared visually and by computer-assisted gel analysis with the type strains for USA300 and USA400. We defined CA-MRSA as MRSA isolates in either the USA300 or the USA400 pulsetypes. Multiplex PCR was performed, as previously described, to type the SCCmec A (29) and to detect the PVL genes (30).
Epidemiologic Data Collection
Epidemiologic data on the isolates were obtained from UHL. These data were age, sex, race/ethnicity, inpatient status, intensive care unit status, long-term-care facility status, hospital admission date, specimen type, specimen collection date, the hospital code number, and the Iowa Reporting Region. Isolates were considered to have been acquired nosocomially if the specimen culture date minus the admission date was >2 days.
Statistical Methods
PFGE patterns and antimicrobial drug susceptibility test results were merged with the demographic data. These data were analyzed with SAS version 9.1 (SAS Institute, Cary, NC, USA) to assess trends in the frequency of USA300/400 in Iowa and to identify possible risk factors for invasive infections with these strains. We used χ2 and adjusted χ2 tests to analyze categorical data and linear regression and logistic regression to analyze continuous data. Alpha was set at 0.05 and all reported p values were 2-tailed.
Seasonality of infections was analyzed by χ2 analysis. Winter was defined as December 22 to March 19, spring as March 20 to June 20, summer as June 21 to September 22, and fall as September 23 to December 21.
The relationships between CA-MRSA and potential risk factors were assessed by univariate analysis. Subsequently, stepwise logistic regression was used to identify factors independently associated with invasive CA-MRSA infection.
Patients infected by USA300/400 MRSA were younger than those infected by other strains (p<0.0001 for both time periods; Tables 1, 2). During 2006, more males than females were infected with USA300/400 (p = 0.06). Most isolates during both time periods were obtained from blood cultures and the distribution of strains did not vary by body site. Most patients were hospitalized for their infections and the proportion of patients admitted to intensive care units did not vary by strain (p = 0.27 and p = 0.35). However, the proportion of MRSA infections that met the definition of nosocomial decreased significantly from 26.1% from 1999–2005 to 16.6% in 2006 (p = 0.0003). During 2006, patients infected with other MRSA strains were more likely than those infected with USA300/400 to have infections that met the definition of nosocomial (p = 0.0006).
Figure 1
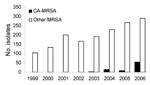
Figure 1. . . . Number of invasive methicillin-resistant Staphylococcus aureus isolates submitted in Iowa, USA, 1999–2006. CA-MRSA, community-associated MRSA.
The antimicrobial susceptibility of 54 invasive USA300/400 isolates is shown in Table 3. None of the USA300 or USA400 isolates had a multidrug-resistant phenotype (e.g., all were resistant to <3 non–β-lactam classes). Specifically, none of the USA300 isolates from Iowa demonstrated the multidrug-resistance pattern described by Diep et al. that is mediated by the multidrug-resistance plasmid pUSA03 (31). All isolates were typeable when the DNA was digested with SmaI. We did not identify any invasive MRSA infections caused by USA300/400 between 1999 and 2002. USA300 caused 3 (1.5%) of 195 infections in 2003, 14 (5.8%) of 243 infections in 2004, 7 (2.5%) of 275 infections in 2005, and 51 (14.9%) of 343 infections in 2006. USA400 caused 2 (0.7%) of 275 infections in 2005 and 3 (0.9%) of 343 infections in 2006. The proportion of MRSA isolates from invasive infections that were CA-MRSA (either USA300 or USA400) increased significantly from 1999–2005 to 2006 (p<0.0001; Figure 1).
Figure 2
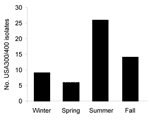
Figure 2. . . . Number of USA300/400 methicillin-resistant Staphylococcus aureus (MRSA) isolates submitted by season, Iowa, USA, 2006.
Reporting region 4, which had the third largest population of the 10 regions, submitted the most isolates; region 1, which had the fourth smallest population, submitted the fewest isolates. We did not find significant differences between the type of MRSA causing infections and the reporting region during 2006. Incidence of infections caused by CA-MRSA varied by season during 2006 (p = 0.0004); a total of 47.3% of these infections occurred during the summer (Figure 2).
The full model for predicting invasive infection with CA-MRSA compared with HA-MRSA included age (young vs. old), seasonality, hospital exposure, and specimen type. However, the only significant predictors of CA-MRSA infection compared with HA-MRSA were age <69 years, which was associated with increased risk (odds ratio [OR] 5.1, 95% confidence interval [CI] 2.06–12.64), and hospital exposure (OR 0.07, 95% CI 0.01–0.51), which was associated with decreased risk.
The current study was unique because it evaluated invasive MRSA isolates from a statewide surveillance system in a rural area. Most prior studies of the epidemiology of CA-MRSA have focused on SSTI among patients in urban areas.
The published literature documents that incidence of CA-MRSA has increased over time in large urban areas. For example, Kaplan et al. found that incidence of CA-MRSA at Texas Children’s Hospital increased each year from August 1, 2001 to July 31, 2004 (32). The percentage caused by USA300 increased from ≈50% in 2000 to >90% in 2003. Of these infections, 95.6% were SSTI and 4.4% were invasive. EMERGEncy ID NET reported that USA300 caused 97% of the MRSA SSTIs seen in emergency rooms in 11 US metropolitan areas during August, 2004 (15). Seybold et al. demonstrated that by 2004 USA300 had become a common cause of MRSA healthcare-associated bloodstream infections (28%) and of nosocomial MRSA bloodstream infections (20%) at Grady Memorial Hospital in Atlanta (21). In contrast, the number of CA-MRSA (primarily USA300) isolates causing invasive infections did not increase substantially in Iowa until 2006.
Klevens et al. published a study of 8,987 invasive MRSA infections reported from the 9 sites in the Active Bacterial Core surveillance (ABCs/Emerging Infections Program Network) from July 2004 through December 2005 (22). The investigators conducted PFGE on 864 (11.3%) of the 7,648 isolates submitted from 8 sites. Of these isolates, 29% were USA300 (16% of the healthcare-associated, hospital-onset infections, 22% of the healthcare-associated, community-onset infections, and 67% of the community-associated, community-onset infections); <0.1% were USA400. In our study, 4.5% of all isolates we typed and 14.9% of isolates from 2006 were USA300, which suggested that the incidence of invasive infections caused by USA300 remains lower in Iowa than in the urban centers studied by Klevens et al.
Unlike the findings of Seybold et al. (21) and Klevens et al (22) from studies conducted in urban areas, USA300/400 rarely caused invasive nosocomial infections in Iowa, a rural state, during the study period. However, unpublished data from the University of Iowa Hospitals and Clinics indicate that these strains are becoming more common causes of invasive nosocomial infections, suggesting that the epidemiology of MRSA may be changing more slowly in rural areas than in large urban areas.
Diep et al. published a follow-up study of previous observations by Han et al. (23) about multidrug-resistant USA300 isolates (31). These investigators found multidrug-resistant isolates in Boston and in San Francisco and they identified male to male sex, past MRSA infection, and use of clindamycin to be risk factors for multiresistant USA300. A multidrug resistance plasmid (pUSA03) mediated these drug resistances. Fortunately, we did not identify this resistance phenotype among our USA300 isolates from Iowa. However, given the rapidity with which plasmid-mediated antimicrobial drug resistance can spread, and given the epidemic nature of USA300, we will continue surveillance for this and other antimicrobial resistances among USA300 isolates in Iowa.
Investigators in the Netherlands, Denmark, and Canada have found nontypeable MRSA among swine (33–37) and persons caring for swine (33–36). Strain ST398, which is not typeable by PFGE after DNA is digested with SmaI, has been found in all of these countries. Iowa produces ≈25% of the swine in the United States. However, we did not identify this strain among the invasive MRSA isolates submitted to the UHL.
Our data did not include information about preceding influenza infections, but we noted that CA-MRSA was isolated twice from the pleural space; other strains of MRSA were not isolated from this site. This finding suggests that CA-MRSA may have caused serious pulmonary infections in a few persons in Iowa. CA-MRSA, particularly USA300, has caused severe infections after influenza (or influenza-like) infections (20,24). During the influenza pandemic of 1918, Chickering and Park noted that many patients acquired severe secondary S. aureus pneumonias following influenza infections (38). Their observations suggest that coincident outbreaks of pandemic influenza and USA300 pneumonia could be catastrophic.
CA-MRSA, particularly USA300, is a public health concern for several other reasons. As noted previously, USA300 is rapidly replacing other strains of MRSA in the community (15,31) and has become an important nosocomial pathogen (21). Moreover, the types of infections caused by USA300 and the epidemiology of this strain are changing rapidly (39). Currently, most USA300 and USA400 isolates have fewer co-resistances than do HA-MRSA isolates (13). However, selective pressures can cause genetic drift in favor of more resistances; papers by Han et al. (23) and Diep et al. (31) documented that USA300 can acquire additional drug resistance determinants. If USA300 and USA400 become resistant to oral antimicrobial agents and the proportion of invasive MRSA infections caused by CA-MRSA continues to increase, many more patients will need parenteral vancomycin therapy, which will increase the difficulty and cost of treating these infections. Furthermore, as the incidence of CA-MRSA infections increases, horizontal transmission of these strains could increase in hospitals, making control of MRSA in healthcare settings even more difficult (6,40). Nosocomial bloodstream infections, ventilator-associated pneumonia, and surgical site infections caused by these strains could be devastating given the necrotizing nature of the infections.
Our study had several limitations. First, the surveillance system was passive. Consequently, demographic data and data about race/ethnicity and exposure to long-term-care facilities were incomplete and data about prior antimicrobial drug exposure and underlying diseases were not available. Additionally, we could identify the region where the specimen originated but not the specific city or county. Moreover, a validation study in Iowa found that hospital laboratories submit only 37% of the invasive MRSA isolates that they identify (D. Dufficy, pers. comm.). However, underreporting affected all regions equally. Furthermore, we used different selection criteria for typing invasive MRSA isolates submitted from 1999–2005 than we did for those submitted in 2006. We typed isolates from 1999–2005 only if they had <3 non–β-lactam coresistances, but we typed all invasive MRSA isolates from 2006. Thus, we may have introduced selection bias that would predispose the incidence of USA300/400 during 1999–2005 toward the null hypothesis (i.e., the annual proportion of MRSA isolates that were USA300/400 did not change during1999 to 2005) but away from the null hypothesis for the incidence of USA300/400 isolates during 1999–2006 (i.e., H0: The annual proportion of MRSA isolates that were USA300/400 was the same in 1999–2005 and in 2006). However, given that all the USA300/400 isolates identified during 2006 would have been detected using the 1999–2005 coresistance selection criterion, we believe that we typed all of the invasive USA300/400 isolates obtained during 1999–2005.
Some might argue that the incidence of invasive infections caused by CA-MRSA increased artificially because physicians were more aware of these organisms in 2006 than they were previously. CA-MRSA certainly has become a hot topic. CA-MRSA was initially identified in the mid 1990s, and many articles about these organisms have been published since then. However, the incidence of invasive infections caused by CA-MRSA in Iowa did not begin rising until 2006. Moreover, Iowa requires laboratories to send all invasive MRSA isolates to the UHL, and laboratory personnel are unlikely to know the details of each patient’s infection. Thus, laboratories probably would not submit isolates of 1 strain preferentially. Finally, many hospitals submitted isolates to the UHL, which suggests that submission bias was unlikely in 2006.
In conclusion, the number of invasive MRSA infections reported in Iowa and the number of invasive infections caused by CA-MRSA increased in Iowa from 1999–2005 to 2006. The increase of CA-MRSA (particularly USA300) poses a unique public health threat. Our study demonstrated that CA-MRSA no longer causes only SSTIs but now causes an increased proportion of invasive infections in a rural state. This finding is particularly disconcerting in light of the severity of these infections and the reports of necrotizing pneumonia caused by USA300 after influenza or influenza-like illness. Surveillance systems must continue to monitor the number and incidence of infections caused by USA300 and to monitor these isolates for changes in antimicrobial susceptibility. The relationship between seasons and CA-MRSA warrants further study.
Mr Van De Griend has an MPH degree from the University of Iowa College of Public Health and is currently a medical student at the University of Iowa Carver College of Medicine. His research interest is molecular epidemiology of staphylococcal infections.
Acknowledgments
We thank Patricia Quinlisk and Deborah L. Dufficy for the work performed by the Iowa Department of Public Health to create and validate the surveillance system. We also thank Michael Pentella for facilitating this study and for critiquing the manuscript. Finally, we thank Richard Hollis, Linda Boyken, Sam Messer, Jennifer Kroeger, and Cassie Dohrn for their assistance with the laboratory methods used in this study.
This study was funded in part by a grant from the Centers for Disease Control and Prevention and was approved by the institutional review board of the University of Iowa College of Medicine.
References
- Barber M. Methicillin-resistant staphylococci. J Clin Pathol. 1961;14:385–93. DOIPubMedGoogle Scholar
- Marshall C, Wesselingh S, McDonald M, Spelman D. Control of endemic MRSA—what is the evidence? A personal view. J Hosp Infect. 2004;56:253–68. DOIPubMedGoogle Scholar
- Diekema DJ, Pfaller MA, Schmitz FJ, Smayevsky J, Bell J, Jones RN, Survey of infections due to Staphylococcus species: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe, and the western Pacific region for the SENTRY antimicrobial surveillance program, 1997–1999. Clin Infect Dis. 2001;32(Suppl 2):S114–32. DOIPubMedGoogle Scholar
- Herold BC, Immergluck LC, Maranan MC, Lauderdale DS, Gaskin RE, Boyle-Vavra S, Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. JAMA. 1998;279:593–8. DOIPubMedGoogle Scholar
- Chambers HF. The changing epidemiology of Staphylococcus aureus? Emerg Infect Dis. 2001;7:178–82. DOIPubMedGoogle Scholar
- Hidron AI, Kourbatova EV, Halvosa JS, Terrell BJ, McDougal LK, Tenover FC, Risk factors for colonization with methicillin-resistant Staphylococcus aureus (MRSA) in patients admitted to an urban hospital: emergence of community-associated MRSA nasal carriage. Clin Infect Dis. 2005;41:159–66. DOIPubMedGoogle Scholar
- Naimi TS, LeDell KH, Como-Sabetti K, Borchardt SM, Boxrud DJ, Etienne J, Comparison of community and health care–associated methicillin-resistant Staphylococcus aureus infection. JAMA. 2003;290:2976–84. DOIPubMedGoogle Scholar
- Vandenesch F, Naimi T, Enright MC, Lina G, Nimmo GR, Heffernan H, Community-acquired methicillin-resistant Staphylococcus aureus carrying Panton-Valentine leukocidin genes: worldwide emergence. Emerg Infect Dis. 2003;9:978–84. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Infectious disease and dermatologic conditions in evacuees and rescue workers after Hurricane Katrina—multiple states, August–September, 2005. MMWR Morb Mortal Wkly Rep. 2005;54:961–4.PubMedGoogle Scholar
- Centers for Disease Control and Prevention. Methicillin-resistant Staphylococcus aureus infections in correctional facilities—Georgia, California, and Texas, 2001–2003. MMWR Morb Mortal Wkly Rep. 2003;52:992–5.PubMedGoogle Scholar
- McDougal LK, Steward CD, Killgore GE, Chaitram JM, McAllister SK, Tenover FC. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: establishing a national database. J Clin Microbiol. 2003;41:5113–20. DOIPubMedGoogle Scholar
- Soderquist B, Berglund C, Stralin K. Community-acquired pneumonia and bacteremia caused by an unusual methicillin-resistant Staphylococcus aureus (MRSA) strain with sequence type 36, staphylococcal cassette chromosome mec type IV and Panton-Valentine leukocidin genes. Eur J Clin Microbiol Infect Dis. 2006;25:604–6. DOIPubMedGoogle Scholar
- Baba T, Takeuchi F, Kuroda M, Yuzawa H, Aoki K, Oguchi A, Genome and virulence determinants of high virulence community-acquired MRSA. Lancet. 2002;359:1819–27. DOIPubMedGoogle Scholar
- Miller LG, Perdreau-Remington F, Rieg G, Mehdi S, Perlroth J, Bayer AS, Necrotizing fasciitis caused by community-associated methicillin-resistant Staphylococcus aureus in Los Angeles. N Engl J Med. 2005;352:1445–53. DOIPubMedGoogle Scholar
- Moran GJ, Krishnadasan A, Gorwitz RJ, Fosheim GE, McDougal LK, Carey RB, Methicillin-resistant S. aureus infections among patients in the emergency department. N Engl J Med. 2006;355:666–74. DOIPubMedGoogle Scholar
- Labandeira-Rey M, Couzon F, Boisset S, Brown EL, Bes M, Benito Y, Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneumonia. Science. 2007;315:1130–3. DOIPubMedGoogle Scholar
- Voyich JM, Otto M, Mathema B, Braughton KR, Whitney AR, Welty D, Is Panton-Valentine leukocidin the major virulence determinant in community-associated methicillin-resistant Staphylococcus aureus disease? J Infect Dis. 2006;194:1761–70. DOIPubMedGoogle Scholar
- Fridkin SK, Hageman JC, Morrison M, Sanza LT, Como-Sabetti K, Jernigan JA, Methicillin-resistant Staphylococcus aureus disease in three communities. N Engl J Med. 2005;352:1436–44. DOIPubMedGoogle Scholar
- Gonzalez BE, Rueda AM, Shelburne SA III, Musher DM, Hamill RJ, Hulten KG. Community-associated strains of methicillin-resistant Staphylococcus aureus as the cause of healthcare-associated infection. Infect Control Hosp Epidemiol. 2006;27:1051–6. DOIPubMedGoogle Scholar
- Hageman JC, Uyeki TM, Francis JS, Jernigan DB, Wheeler JG, Bridges CB, Severe community-acquired pneumonia due to Staphylococcus aureus, 2003–04 influenza season. Emerg Infect Dis. 2006;12:894–9. DOIPubMedGoogle Scholar
- Seybold U, Kourbatova EV, Johnson JG, Halvosa SJ, Wang YF, King MD, Emergence of community-associated methicillin-resistant Staphylococcus aureus USA300 genotype as a major cause of health care–associated blood stream infections. Clin Infect Dis. 2006;42:647–56. DOIPubMedGoogle Scholar
- Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA. 2007;298:1763–71. DOIPubMedGoogle Scholar
- Han LL, McDougal LK, Gorwitz RJ, Mayer KH, Patel JB, Sennott JM, High frequencies of clindamycin and tetracycline resistance in methicillin-resistant Staphylococcus aureus pulsed-field type USA300 isolates collected at a Boston ambulatory health center. J Clin Microbiol. 2007;45:1350–2. DOIPubMedGoogle Scholar
- Francis JS, Doherty MC, Lopatin U, Johnston CP, Sinha G, Ross T, Severe community-onset pneumonia in healthy adults caused by methicillin-resistant Staphylococcus aureus carrying the Panton-Valentine leukocidin genes. Clin Infect Dis. 2005;40:100–7. DOIPubMedGoogle Scholar
- Iowa Antibiotic Resistance Surveillance Program [homepage on the Internet]. 2006 [cited 2009 Jul 24]. Available from http://www.uhi.uiowa.edu/services/antibioticresistance/index.xml
- Diseases reportable to the Iowa Department of Public Health [homepage on the Internet]. 2004 [cited 2008 Jul 24]. Available from http://www.idph.state.ia.us/hcci/common/pdf/naeyc/ia_reportable_diseases.pdf
- Clinical and Laboratory Standards Institute [homepage on the Internet]. 2006 [cited 2008 Jul 24]. Available from http://www.clsi.org
- Diekema DJ, Pfaller MA, Turnidge J, Verhoef J, Bell J, Fluit AC, Genetic relatedness of multidrug-resistant, methicillin (oxacillin)-resistant Staphylococcus aureus bloodstream isolates from SENTRY antimicrobial resistance surveillance centers worldwide, 1998. Microb Drug Resist. 2000;6:213–21. DOIPubMedGoogle Scholar
- Oliveira DC. Lencastre Hd. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2002;46:2155–61. DOIPubMedGoogle Scholar
- Lina G, Piemont Y, Godail-Gamot F, Bes M, Peter MO, Gauduchon V, Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. 1999;29:1128–32. DOIPubMedGoogle Scholar
- Diep BA, Chambers HF, Graber CJ, Szumowski JD, Miller LG, Han LL, Emergence of multidrug-resistant, community-associated, methicillin-resistant Staphylococcus aureus clone USA300 in men who have sex with men. Ann Intern Med. 2008;148:249–57.PubMedGoogle Scholar
- Kaplan SL, Hulten KG, Gonzalez BE, Hammerman WA, Lamberth L, Versalovic J, Three-year surveillance of community-acquired Staphylococcus aureus infections in children. Clin Infect Dis. 2005;40:1785–91. DOIPubMedGoogle Scholar
- de Neeling AJ, van den Broek MJ, Spalburg EC, van Santen-Verheuvel MG, Dam-Deisz WD, Boshuizen HC, High prevalence of methicillin resistant Staphylococcus aureus in pigs. Vet Microbiol. 2007;122:366–72. DOIPubMedGoogle Scholar
- Armand-Lefevre L, Ruimy R, Andremont A. Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg Infect Dis. 2005;11:711–4. DOIPubMedGoogle Scholar
- Voss A, Loeffen F, Bakker J, Klaassen C, Wulf M. Methicillin-resistant Staphylococcus aureus in pig farming. Emerg Infect Dis. 2005;11:1965–6. DOIPubMedGoogle Scholar
- Khanna T, Friendship R, Dewey C, Weese JS. Methicillin resistant Staphylococcus aureus colonization in pigs and pig farmers. Vet Microbiol. 2007;128:298–03. DOIPubMedGoogle Scholar
- Guardabassi L, Stegger M, Skov R. Retrospective detection of methicillin resistant and susceptible Staphylococcus aureus ST398 in Danish slaughter pigs. Vet Microbiol. 2007;122:384–6. DOIPubMedGoogle Scholar
- Ellis MW, Hospenthal DR, Dooley DP, Gray PJ, Murray CK. Natural history of community-acquired methicillin-resistant Staphylococcus aureus colonization and infection in soldiers. Clin Infect Dis. 2004;39:971–9. DOIPubMedGoogle Scholar
- Lescure FX, Biendo M, Douadi Y, Schmit JL, Eveillard M. Changing epidemiology of methicillin-resistant Staphylococcus aureus and effects on cross-transmission in a teaching hospital. Eur J Clin Microbiol Infect Dis. 2006;25:205–7. DOIPubMedGoogle Scholar
Figures
Tables
Follow Up
Earning CME Credit
To obtain credit, you should first read the journal article. After reading the article, you should be able to answer the following, related, multiple-choice questions. To complete the questions and earn continuing medical education (CME) credit, please go to http://www.medscape.com/cme/eid. Credit cannot be obtained for tests completed on paper, although you may use the worksheet below to keep a record of your answers. You must be a registered user on Medscape.com. If you are not registered on Medscape.com, please click on the New Users: Free Registration link on the left hand side of the website to register. Only one answer is correct for each question. Once you successfully answer all post-test questions you will be able to view and/or print your certificate. For questions regarding the content of this activity, contact the accredited provider, CME@medscape.net. For technical assistance, contact CME@webmd.net. American Medical Association’s Physician’s Recognition Award (AMA PRA) credits are accepted in the US as evidence of participation in CME activities. For further information on this award, please refer to http://www.ama-assn.org/ama/pub/category/2922.html. The AMA has determined that physicians not licensed in the US who participate in this CME activity are eligible for AMA PRA Category 1 Credits™. Through agreements that the AMA has made with agencies in some countries, AMA PRA credit is acceptable as evidence of participation in CME activities. If you are not licensed in the US and want to obtain an AMA PRA CME credit, please complete the questions online, print the certificate and present it to your national medical association.
Article Title: Community-Associated Methicillin-Resistant Staphylococcus aureus, Iowa, USA
CME Questions
1. A 25-year-old man is admitted to the intensive care unit with head trauma after a motor vehicle accident, receives a urinary catheter, and is ventilated. He receives prophylactic antibiotics for multiple skin wounds. Which of the following is least likely to be a risk factor for healthcare-associated (HA) verus community-associated (CA) methicillin-resistant Staphylococcus aureus (MRSA) in this patient?
A. Young age
B. Hospitalization
C. Catheter use
D. Exposure to antimicrobials
2. Which of the following best describes the definition of multiresistant isolates of MRSA?
A. Resistant to more than 4 of 8 antimicrobial classes
B. Resistant to more than 2 of 7 antimicrobial classes
C. Resistant to more than 3 of 8 antimicrobial classes
D. Resistant to more than 3 of 7 antimicrobial classes
3. Which of the following most accurately describes the trend in CA-MRSA USA300/400 infection in 1 US state from 1999 to 2006?
A. Most USA300/400 isolates were from the skin
B. Distribution of MRSA strains varied according to body site
C. The rate of nosocomial infections increased significantly
D. USA300/400 isolates did not demonstrate multidrug resistance
4. A 64-year-old man in a long-term care facility is admitted to the hospital in the summer for acute stroke. Which of the following is most likely to be a significant positive predictor of CA-MRSA compared with HA-MRSA in this patient?
A. Admission in summer versus other season
B. Age younger than 69 years
C. Hospitalization
D. Concurrent urinary tract infection
5. Which of the following best describes recent patterns of CA-MRSA infection in the United States?
A. Increase in incidence of USA300 strain of MRSA
B. Over 50% of CA-MRSA infections are invasive
C. USA300 is associated with 60% of MRSA bloodstream infections
D. Decrease in urban areas and increase in rural areas
Activity Evaluation
|
1. The activity supported the learning objectives. |
||||
|
Strongly Disagree |
|
|
|
Strongly Agree |
|
1 |
2 |
3 |
4 |
5 |
|
2. The material was organized clearly for learning to occur. |
||||
|
Strongly Disagree |
|
|
|
Strongly Agree |
|
1 |
2 |
3 |
4 |
5 |
|
3. The content learned from this activity will impact my practice. |
||||
|
Strongly Disagree |
|
|
|
Strongly Agree |
|
1 |
2 |
3 |
4 |
5 |
|
4. The activity was presented objectively and free of commercial bias. |
||||
|
Strongly Disagree |
|
|
|
Strongly Agree |
|
1 |
2 |
3 |
4 |
5 |
Related Links
Table of Contents – Volume 15, Number 10—October 2009
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Loreen A. Herwaldt, 200 Hawkins Dr, Iowa City, IA 52242–1081, USA
Top