Volume 8, Number 1—January 2002
Dispatch
Presence of Rickettsia felis in the Cat Flea from Southwestern Europe1
Abstract
Rickettsia felis, formerly called ELB agent, was identified by using molecular biology techniques in the cat flea (Ctenocephalides felis felis) from southwestern Spain. For the first time this flea-transmitted rickettsia has been detected within its vector in Eurasia.
Members of the genus Rickettsia are commonly associated with hematophagous arthropods such as ticks, fleas, or lice. Rickettsia felis, formerly ELB agent, was detected in 1990 when tissues from the cat flea, Ctenocephalides felis, were examined under electron microscopy. After this, several antigenic and molecular studies concerning this rickettsia were developed (1). R. felis is maintained in cat fleas by transovarian transmission (2). Infection in humans has been described in the USA (3), Mexico (4), and Brazil (1) by polymerase chain reaction (PCR) amplification and recently in France by serologic tests (1).
During a study concerning rickettsial organisms transmitted by ticks in southwest Spain, using molecular tools for diagnosis, a rickettsial microorganism was detected in some cat fleas on domestic cats and dogs from different counties of the Cadiz Province.
Figure
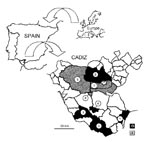
Figure. Localization of randomly selected sampling points in Cadiz Province for cat fleas with indication of the presence of Rickettsia felis by polymerase chain reaction procedures. Localities are 1. San Roque, 2....
The fleas used in this study (60 females and 11 males) were collected, together with ticks, from 2 cats and 12 dogs from eight localities of Cadiz Province in southwestern Spain from May to August of 1999 (Figure, Table). The hosts were domestic and peridomesticated dogs and cats living in a range of health-care conditions. Collected fleas were fixed in 70% ethyl alcohol and stored at 4°C until they were processed. Taxonomic determination was made by using current taxonomic keys (5,6). All specimens subjected to analysis were C. felis felis (Bouché, 1835).
DNA was extracted from 14 lots of fleas (ranging from 1 to 11 specimens per lot) by using the DNeasy_Tissue kit(Qiagen GmbH, Hilden, Germany) (7). Elution of DNA was made in 100 μL of TE buffer (1 mM Tris HCl, 0.1 mM EDTA). Extraction blanks, consisting of water processed along with flea samples, were also included as controls.
A Biometra DNA Thermalcycler (Gottingen, Germany) was used for all PCR amplification. Three microliters of each DNA extraction were added to 27 μL of master mixture for each reaction. Final reagent concentration was 0.2 μM for each primer, 200 μM for each deoxynucleotide triphosphate (Promega Corp., Madison, WI), 2 U of Biotaq polymerase (BioLine, London, UK), and 1x Bioline buffer. The following thermal cycler parameters were used with the primer pairs for citrate synthase (glta) RpCS.877p and RpCS1258n (8), 120-kDa genus common antigen (ompB) (rfompbf: 5’–GAC AAT TAA TAT CGG TGA CGG, and rfompbr: 5’-TGC ATC AGC ATT ACC GCT TGC), 190-kDa protein antigen (ompA) Rr190.70p, and Rr190.602n (8): 96°C (90 sec), followed by 35 cycles of 94°C (30 sec), 50°C (30 sec), and 72°C (45 sec), followed by an extension period (72°C, 7 min). For the amplification of a 426-base pair fragment of 16S rRNA gene, we used the primers fD1 (9) and Rc16S.452n (10) and 59°C as annealing temperature.
Seven lots from five localities around Cadiz and Gibraltar bays were positive to amplification of fragments of 16S rRNA, glta, ompA and ompB genes.
In brief, after amplification, primers and nucleotides were removed from 300 μL of PCR products by purification on the Wizard PCR preps purification system (Promega, Madison, WI). Approximately 100 fmol of the purified PCR product (4-5 μL) were used directly in the sequencing reaction.
The PCR cycle sequencing was performed for each amplicon by using the correct forward or reverse primers and the Silver sequence DNA Sequencing System (Promega). Sequencing reaction products were loaded twice on 40 cm 6% polyacrylamide 7M urea gels by electrophoresis in the Sequi-Gen Nucleic Acid Sequencing System (BioRad, Hercules, CA) at 55 W of constant electrophoresis (55°C) and separated for 4 hr 30 min. and 2 hr 30 min, respectively. Gel was silver stained by using the standard Promega protocol. A permanent record was made in scanning the gel. To determine the sequence of positions near primers, we used a 10% polyacrylamide 7M urea electrophoresis gel. The sequence of both strings was determined twice for each fragment.
Sequences obtained were compared with those from other Rickettsia species in GenBank by using the BLAST utility (National Center for Biotechnology Information, Bethesda, MD) and FASTA routine from GCG environment. Fragment sequence for 16S rRNA, glta, ompA, and ompB sequence were identical to previously reported sequence for R. felis. The 16S rRNA amplified fragment was identical to previously reported sequence (GenBank L28944) between positions 1 and 410 (3). The fragment sequenced for citrate synthase corresponded to positions 757 and 1138 in GenBank accession AF210692 (1). The fragment amplified for ompA corresponded to positions 478 to 987 in GenBank accession AF191026 (11). The fragment amplified for ompB corresponded to positions 599 to 1259 in GenBank accession AF210695 (1). Amplification was unsuccessful in all negative controls.
R. felis has been found extensively in commercial colonies and natural cat fleas, parasitizing a large range of mammalian hosts in several states of the United States (12,13).
For the first time R. felis was detected in Eurasia, by means of PCR and partial sequencing of genes classically used in rickettsial molecular characterization and phylogeny. The sequences of glta, ompA, ompB, and 16S rRNA from Cadiz cat fleas were identical to the homologous sequences previously reported for R. felis obtained from fleas reared in EL Laboratories (Soquel, CA) (3) and Louisiana State University (11) and isolated by Flea Data Inc. (Freeville, NY) (1).
In humans, R. felis may produce a clinical syndrome similar to murine typhus (3). Thus, R. felis could be implicated in murine typhus-compatible cases detected in southwest Spain (14), especially since the oriental rat flea, Xenopsylla cheopis (Rothschild, 1903), is absent from this area.
Thirteen species of flea belonging to the genus Ctenocephalides have been described to date (15), mainly distributed in continental Africa (16), with a worldwide contemporary distribution in a large range of hosts, mainly anthropic species of the group (C. felis), which has a large potential host range. The primary source of the bacterium might be Africa, where this flea genus apparently originated.
This work was supported in part by grants from the Fondo de Investigación Sanitaria (FIS-99/0296), Servicio Andaluz de Salud (SAS-203/98) of Junta de Andalucía, and Universidad de Jaén (UJA-FLV/jbf) research programs.
Dr. Márquez is associate professor of zoology and parasitology at the Faculty of Experimental Sciences at Jaén University, Spain. His research interests focus on medical entomology and arthropod-borne diseases.
References
- Raoult D, La Scola B, Enea M, Fournier PE, Roux V, Fenollar F, A flea-associated Rickettsia pathogenic for humans. Emerg Infect Dis. 2001;7:73–81.PubMedGoogle Scholar
- Adams JR, Schmidtmann ET, Azad AF. Infection of colonized cat fleas, Ctenocephalides felis (Bouché), with a rickettsia-like microorganism. Am J Trop Med Hyg. 1990;43:400–9.PubMedGoogle Scholar
- Schriefer ME, Sacci JB Jr, Dumler JS, Bullen MG, Azad AF. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J Clin Microbiol. 1994;32:949–54.PubMedGoogle Scholar
- Zavala-Velázquez JE, Ruíz-Sosa JA, Sánchez-Elias RA, Becerra-Carmona G, Walker DH. Rickettsia felis rickettsiosis in Yucatán. Lancet. 2000;356:1079–80. DOIPubMedGoogle Scholar
- Beaucournu JC, Launay H. Les puces de France et du bassin méditerranéen occidental. Faune de France 76. Paris: Féderation Française des Sociétés de Sciences Naturelles; 1990.
- Ménier K, Beaucournu JC. Taxonomic study of the genus Ctenocephalides Stiles & Collins, 1930 (Insecta: Siphonaptera: Pulicidae) by using aedeagus characters. J Med Entomol. 1998;35:883–90.PubMedGoogle Scholar
- Dneasy Tissue Kit Handbook. Hilden, Germany: Quiagen GmbH; version April 1999. #1011607.
- Regnery RL, Spruill CL, Plikaytis BD. Genotypic identification of rickettsiae and stimation of intraspecies sequence divergence for portions of two rickettsial genes. J Bacteriol. 1991;173:1576–89.PubMedGoogle Scholar
- Weisburg WG, Dobson ME, Samuel JE, Dasch GA, Mallavia L, Baca O, Phylogenetic diversity of the rickettsiae. J Bacteriol. 1989;171:4302–6.
- Márquez FJ, Muniain MA, Soriguer RC, Izquierdo G, Rodríguez-Baño J, Borobio MV. Genotypic identification of an undescribed spotted fever group Rickettsia in Ixodes ricinus from Southwestern Spain. Am J Trop Med Hyg. 1998;58:570–7.PubMedGoogle Scholar
- Bouyer DH, Stenos J, Croquet-Valdes PA, Foil LD, Walker DH. The identification and characterization of a previously undiscovered rOmpA-encoding gene in Rickettsia felis. In: Raoult D, Brouqui P, editors. Rickettsia and rickettsial diseases at the turn of the third millennium. Marseille: Elsevier; 1999. p. 11-15.
- Azad AF, Radulovic S, Higgins JA, Noden BH, Troyer JM. Flea-borne rickettsiosis: ecologic considerations. Emerg Infect Dis. 1997;3:319–27.PubMedGoogle Scholar
- Higgins JA, Sacci JB, Schriefer ME, Endris RG, Azad AF. Molecular identification of rickettsia-like microorganisms associated with colonized cat fleas (Ctenocephalides felis). Insect Mol Biol. 1994;3:27–33.PubMedGoogle Scholar
- Bernabeu-Wittel M, Pachón J, Alarcón A, López-Cortés LF, Viciana P, Jimenez-Mejias ME, Murine typhus as a common cause of fever of intermediate duration. A 17-year study in the South of Spain. Arch Intern Med. 1999;159:872–6. DOIPubMedGoogle Scholar
- Beaucournu JC, Ménier K. Le genre Ctenocephalides Stiles et Collins, 1930 (Siphonaptera, Pulicidae). Parasite. 1998;5:3–16.PubMedGoogle Scholar
- Ménier K, Beaucournu JC. Approche biogeographique du genre Ctenocephalides Stiles et Collins, 1930 (Insecta: Siphonaptera). Biogeographica. 1999;75:79–88.
Figure
Table
Cite This Article1This study was presented in part at the II National Meeting of the Rickettsia and Borrelia Group (Spanish Society of Infectious Diseases and Clinical Microbiology), Sitges, Barcelona, Spain, October 2000.
Table of Contents – Volume 8, Number 1—January 2002
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Francisco J. Márquez, Departamento de Biología Animal, Biología Vegetal y Ecología, Universidad de Jaén, Paraje Las Lagunillas s/n, 23071 Jaén, Spain; fax: 34-53-012141;
Top