Volume 10, Number 3—March 2004
Dispatch
Endemic Carbapenem-resistant Pseudomonas aeruginosa with Acquired Metallo-β-lactamase Determinants in European Hospital
Abstract
Acquired metallo-β-lactamases (MBLs) can confer broad-spectrum β-lactam resistance (including carbapenems) not reversible by conventional β-lactamase inhibitors and are emerging resistance determinants of remarkable clinical importance. In 2001, multidrug-resistant Pseudomonas aeruginosa carrying blaVIM MBL genes were found to be widespread (approximately 20% of all P. aeruginosa isolates and 70% of the carbapenem-resistant isolates) at Trieste University Hospital. Clonal diversity and heterogeneity of resistance determinants (either blaVIM-1-like or blaVIM-2-like) were detected among MBL producers. This evidence is the first that acquired MBLs can rapidly emerge and establish a condition of endemicity in certain epidemiologic settings.
Bacterial pathogens bearing acquired metallo-β-lactamase (MBL) genes exhibit a broad-spectrum resistance to β-lactams that is not reversible by serine-β-lactamase inhibitors (e.g., clavulanate and penicillanic acid sulphones), since MBLs are capable of hydrolyzing most β-lactams and are not susceptible to inhibitors. Because of the efficient carbapenemase activity of these enzymes, the resistance profile of MBL producers notably includes also carbapenems, which are the β-lactams with the broadest spectrum of activity and are among the “last resort” drugs for the treatment of gram-negative nosocomial infections. In addition, MBL producers most often exhibit resistant phenotype to additional classes of drugs since they originate nosocomially and acquired MBL genes typically cluster with other drug resistance determinants in the variable region of multi-resistance integrons (1–3). For these reasons, infections caused by MBL producers can pose a substantial challenge for antimicrobial chemotherapy.
The IMP and VIM enzymes are the most common types of acquired MBLs (2,3). The IMP enzymes were first reported in Japan (4), while the VIM enzymes were first reported in Europe (5), but both types of enzymes are now emerging in Asia, Europe, and the Americas as acquired resistance determinants in nosocomial isolates of Enterobacteriaceae, Pseudomonas aeruginosa, Acinetobacter spp. and other nonfastidious, gram-negative nonfermenters (3). The VIM-1 enzyme is 90% amino acid homologous with the VIM-2 variant and <40% amino acid homologous with the IMP enzymes (3). Both types of resistance genes are carried on mobile gene cassettes inserted into plasmid- or chromosomal-borne integrons, a location that eventually facilitates horizontal spreading among different strains (3).
Thus far, strains with acquired MBLs have usually been reported sporadically or as causing small nosocomial outbreaks (4,6–8), while longitudinal surveys have demonstrated, at most, a low-level endemicity of MBL producers in hospitals where similar strains have been detected (9,10). One major hospital outbreak, caused by an MBL-producing P. aeruginosa clone, was recently reported in Greece (11). We describe the emergence of high-level-endemicity for MBL-producing P. aeruginosa, which has recently occurred in a hospital setting of southern Europe.
In the University Hospital of Trieste (northern Italy, at the border with Slovenia), clinical isolates of P. aeruginosa producing VIM-type MBLs were detected sporadically, for the first time, in 1999 (12). In 2001, a significant increase in the prevalence of imipenem-resistant P. aeruginosa isolates was observed at the Laboratory of Clinical Microbiology of that hospital (29%, vs. 19% in 2000 and 21% in 1999, respectively; p < 0.001 according to the χ2 test; statistical analyses were conducted with Epi Info statistical software, version 6.03, Centers for Disease Control and Prevention, Atlanta, GA).
Of the 444 nonreplicate imipenem-resistant P. aeruginosa isolates collected in 2001, a total of 89 were randomly selected and analyzed for acquired MBL genes of the blaIMP and blaVIM types in dot-blot hybridization experiments carried out with purified genomic DNA spotted (0.5 μg per spot) on positively charged nylon membranes (ZetaProbe, Bio-Rad, Hercules, CA) with digoxygenin-labeled DNA probes. The probes were polymerase chain reaction amplicons containing internal fragments of the blaIMP-1 (754–1,114 nt, EMBL/GenBank database entry S71932) or of the blaVIM-1 gene (3,366–3,888 nt, EMBL/GenBank database entry Y18050), respectively obtained using primers IMP-DIA (forward, 5′-GGAATAGAGTGGCTTAATTCTC; reverse, 5′-GTGATGCGTCYCCAAYTTCACT) and VIM-DIA (forward, 5′-CAGATTGCCGATGGTGTTTGG; reverse, 5′-AGGTGGGCCATTCAGCCAGA) as described previously (13). Hybridization was carried out under conditions that allowed recognition, by each probe, of different allelic variants of the corresponding MBL determinant. None of the imipenem-resistant isolates were recognized by the blaIMP probe, while 64 (72%) were recognized by the blaVIM probe. In the 64 blaVIM-positive isolates, the nature of the determinant was further investigated by analysis of the RsaI restriction fragment length polymorphism of the gene region amplified by the VIM-DIA primers as described previously. With this approach, the determinant was identified as blaVIM-1-like in 54 isolates (84%), and as blaVIM-2-like in the remaining 10 isolates (16%).
Figure
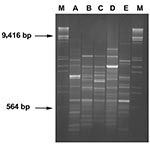
The sources of the 64 blaVIM-positive isolates were 52 inpatients from 15 different wards (including 10 medical wards, 4 surgical wards, and an intensive care unit), 5 patients from 4 different long-term care facilities for elderly persons, and 7 outpatients (Table 1). The degree of genomic relatedness of these isolates was investigated by Random Amplification of Polymorphic DNA (RAPD) (14) and by Amplified Fragment Length Polymorphism (AFLP) (15). Electrophoretic profiles generated by the techniques described earlier were compared by the GelComparII software (Applied Maths, Kortrijk, Belgium). Consistent results were obtained with both typing methods. Isolates sharing a Dice similarity coefficient >0.88 comparing their RAPD-profiles were assigned to the same cluster. Results of molecular typing indicated that most blaVIM-positive isolates (61 [95%]) belonged to either of two clusters, indicated as cluster A and B respectively, while the remaining three isolates were unrelated with those clusters and also among each other (Figure). Cluster A included 53 isolates, all containing blaVIM-1-like determinants. They were widely distributed in the hospital (15 wards), and were also found in three long-term care facilities and in six outpatients. Cluster B included eight isolates, all containing blaVIM-2-like determinants. The isolates were from four wards where isolates of cluster A had also been detected. Of the three sporadic isolates, one (carrying a blaVIM-2-like gene) was from a ward where isolates of clusters A and B had also been detected, the second (also carrying a blaVIM-2-like gene) was from a long-term care facility different from those yielding isolates of cluster A, and the third (carrying a blaVIM-1-like gene) was from an outpatient (Table 1). Genotyping of the 25 blaVIM-negative isolates indicated that 5 belonged in cluster A, 1 in cluster B, while the remaining 19 were unrelated to the VIM producers and were overall distributed among 6 different genotypes (Table 1).
Imipenem MICs for the blaVIM-positive isolates were always >64 μg/mL (range 64–512 μg/mL), while being always <64 μg/mL for the hybridization-negative isolates. Most of the blaVIM-positive isolates (49 of 64 [76%]) exhibited a multidrug-resistant phenotype including all the tested drugs (imipenem, meropenem, ceftazidime, piperacillin, aztreonam, amikacin, gentamicin, tobramycin, and ciprofloxacin), except polymixin B. On the other hand, this virtually panresistant phenotype was observed in 7 (28%) of 25 blaVIM-negative isolates (Table 2).
Our findings are of concern since they demonstrate that acquired MBLs can rapidly emerge and become a major cause of broad-spectrum β-lactam resistance among nosocomial pathogens. In our setting blaVIM-positive P. aeruginosa isolates, which were sporadically detected for the first time in 1999 (12), represented approximately 20% of all P. aeruginosa isolates and 70% of the carbapenem-resistant P. aeruginosa isolates, respectively, during 2001. These figures exceed those reported for MBL producers from other settings (7,9,10). As an additional matter of concern, the blaVIM-positive isolates were significantly more resistant than the blaVIM-negative isolates to non-β-lactam antimicrobial agents as well.
In this survey, the blaVIM-positive isolates were detected on a regular basis during the year and appeared to be widely distributed in the hospital and even outside of it. Molecular characterization showed the simultaneous circulation of different blaVIM alleles (either blaVIM1-like or blaVIM-2-like) in multiple P. aeruginosa clones. Overall, these findings suggest that blaVIM determinants have rapidly established a condition of high-level endemicity in this area. To the best of our knowledge, this study is the first in which a similar condition has been reported. Even the large outbreak reported in Greece was caused by a single clone and was apparently confined to the hospital wards (11). The finding of blaVIM-negative P. aeruginosa isolates showing the same genotype as that of the two major clusters of blaVIM-positive strains suggests a likely acquisition of the MBL determinants by strains already endemic in this area, followed by clonal expansion of the blaVIM-positive strains.
The possibility that spreading transferable MBL genes among nosocomial gram-negative pathogens could emerge as a major problem in the clinical setting underscores the need for systematic surveillance of these resistance determinants. Considering that MBL producers were also isolated from outpatients and from long-term care facility patients, even if all of them showed at least one hospital treatment during the 6 months before, surveillance should not be restricted to nosocomial isolates but should also include isolates from community-acquired infections.
Dr. Lagatolla is a research scientist of the Dipartimento di Scienze Biomediche–Sezione di Microbiologia at the University of Trieste, Italy. Her work focuses on epidemiologic surveillance of nosocomial infections, with particular attention to the spread of the determinants of antimicrobial resistance.
Acknowledgment
This work was supported by grants from Italian M.I.U.R. (nos. 2001068755-005 and 20011068755-003), and by grant no. HPRN-CT-2002-00264 from the European Union (MEBEL project).
References
- Livermore DM, Woodford N. Carbapenemases: a problem in waiting? Curr Opin Microbiol. 2000;3:489–95. DOIPubMedGoogle Scholar
- Bush K. New β-lactamases in gram-negative bacteria: diversity and impact on the selection of antimicrobial therapy. Clin Infect Dis. 2001;32:1085–9. DOIPubMedGoogle Scholar
- Nordmann P, Poirel L. Emerging carbapenemases in Gram-negative aerobes. Clin Microbiol Infect. 2002;8:321–31. DOIPubMedGoogle Scholar
- Osano E, Arakawa Y, Wacharotayankun R, Ohta M, Horii T, Ito H, Molecular characterization of an enterobacterial metallo-β-lactamase found in a clinical isolate of Serratia marcescens that shows imipenem resistance. Antimicrob Agents Chemother. 1994;38:71–8.PubMedGoogle Scholar
- Lauretti L, Riccio ML, Mazzariol A, Cornaglia G, Amicosante G, Fontana R, Cloning and characterization of blaVIM, a new integron-borne metallo-β-lactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob Agents Chemother. 1999;43:1584–90.PubMedGoogle Scholar
- Senda K, Arakawa Y, Nakashima K, Ito I, Ichiyama S, Shimokata K, Multifocal outbreaks of metallo-β-lactamase-producing Pseudomonas aeruginosa resistant to broad-spectrum β-lactams, including carbapenems. Antimicrob Agents Chemother. 1996;40:349–53.PubMedGoogle Scholar
- Senda K, Arakawa Y, Ichiyama S, Nakashima K, Ito H, Ohsuka S, PCR detection of metallo-β-lactamase gene (blaIMP) in gram-negative rods resistant to broad-spectrum β-lactams. J Clin Microbiol. 1996;34:2909–13.PubMedGoogle Scholar
- Cornaglia G, Mazzariol A, Lauretti L, Rossolini GM, Fontana R. Hospital outbreak of carbapenem-resistant Pseudomonas aeruginosa producing VIM-1, a novel transferable metallo-β-lactamase. Clin Infect Dis. 2000;31:1119–25. DOIPubMedGoogle Scholar
- Hirakata Y, Izumikawa K, Yamaguchi T, Takemura H, Tanaka H, Yoshida R, Rapid detection and evaluation of clinical characteristics of emerging multiple-drug-resistant gram-negative rods carrying the metallo-β-lactamase gene blaIMP. Antimicrob Agents Chemother. 1998;42:2006–11.PubMedGoogle Scholar
- Lee K, Lim JB, Yum JH, Yong D, Chong Y, Kim JM, blaVIM-2 cassette-containing novel integrons in metallo-β-lactamase-producing Pseudomonas aeruginosa and Pseudomonas putida isolates disseminated in a Korean hospital. Antimicrob Agents Chemother. 2002;46:1053–8. DOIPubMedGoogle Scholar
- Tsakris A, Pournaras S, Woodford N, Palepou MF, Babini GS, Duoboyas J, Outbreak of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J Clin Microbiol. 2000;38:1290–2.PubMedGoogle Scholar
- Rossolini GM, Riccio ML, Cornaglia G, Pagani L, Lagatolla C, Selan L, Carbapenem-resistant Pseudomonas aeruginosa with acquired blaVIM metallo-β-lactamase determinants, Italy. Emerg Infect Dis. 2000;6:312–3. DOIPubMedGoogle Scholar
- Migliavacca R, Docquier JD, Mugnaioli C, Amicosante G, Daturi R, Lee K, Simple microdilution test for detection of metallo-beta-lactamase production in Pseudomonas aeruginosa. J Clin Microbiol. 2002;40:4388–90. DOIPubMedGoogle Scholar
- Mahenthiralingam E, Campbell ME, Foster J, Lam JS, Speert DP. Random amplified polymorphic DNA typing of Pseudomonas aeruginosa isolates recovered from patients with cystic fibrosis. J Clin Microbiol. 1996;34:1129–35.PubMedGoogle Scholar
- Speijer H, Savelkoul PHM, Bonten MJ, Stobberingh EE, Tjhie JH. Application of different genotyping methods for Pseudomonas aeruginosa in a setting of endemicity in an intensive care unit. J Clin Microbiol. 1999;37:3654–61.PubMedGoogle Scholar
Figure
Tables
Cite This ArticleTable of Contents – Volume 10, Number 3—March 2004
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Cristina Lagatolla, Dipartimento Scienze Biomediche, Università di Trieste, Via Fleming 22, 34127 Trieste, Italy; fax: +39-040-351-668
Top