Volume 13, Number 7—July 2007
Perspective
Brazilian Vaccinia Viruses and Their Origins
Abstract
Although the World Health Organization (WHO) declared global smallpox eradicated in 1980, concerns over emergent poxvirus infections have increased. Most poxvirus infections are zoonotic; exploring their genetic diversity will illuminate the genetic and evolutionary aspects of poxvirus infections, ecology, and epidemiology. In recent decades, several strains of the orthopoxvirus vaccinia virus (VACV) have been isolated throughout Brazil, including many genetically distinct isolates within the same outbreak. To further investigate the diversity and origins of these viruses, we analyzed molecular data from 8 Brazilian VACV isolates and compared several genes involved in virus structure and pathogenicity. Genetic variation among isolates suggests that ancestral Brazilian VACVs existed before the beginning of the WHO smallpox eradication vaccination campaigns and that these viruses continue to circulate.
In 1980, the World Health Organization (WHO), after a massive vaccination program, announced the eradication of smallpox, the contagious and deadly disease caused by Variola virus (VARV). This program used live vaccinia virus (VACV), a virus from the same genus, orthopoxvirus, which shares a high degree of immunologic cross-reactivity with VARV. Recent reports of cowpox virus (CPXV) infections in Europe, monkeypox virus (MPXV) outbreaks in Africa and the United States, and the surprising emergence of VACV in Brazil highlight the need for continued research into the ecology, epidemiology, origin, and evolution of these viruses (1).
Figure 1

The known history of VACV species imported to Brazil dates back to 1804, when human vaccine arrived at a port in Bahia State on the arms of slaves returning to Brazil from Lisbon, Portugal (2) (Figure 1). From Bahia State the slaves were sent south to Rio de Janeiro State, possibly passing the vaccine to local people as they made their way through the region. Indeed, from 1804 to 1887, the Brazilian population (including slaves and other people living in the countryside) was vaccinated in this manner, arm to arm. In some cities, vaccination was obligatory, beginning in 1832. In 1887, the first animal vaccine produced in calves was imported in flasks to the vaccine institute in Rio de Janeiro (now Oswaldo Cruz Institute) from the Chambon Institute in Paris, France (2). The vaccine was then distributed to other states across Brazil, including Minas Gerais, Espirito Santo, São Paulo, Mato Grosso, Rio Grande do Sul, and Pernambuco (2). During this time, between 1887 and 1895, vaccine institutes were established in these states (2).
Published accounts of Brazilian vaccinia-like viruses isolated from sentinel mice and recent outbreaks on dairy farms affecting cattle and their handlers suggest the once-emergent disease has now become endemic (4–13). We combined historical information and published data to develop insights into the possible origin(s) of the Brazilian VACV (BRZ-VACV) now established in Brazil. Our goal was to determine whether BRZ-VACV represents an escaped vaccine strain, an autochthonous orthopoxvirus, or both.
We examined the sequence diversity of 8 geographically and temporally variable BRZ-VACV isolates. We compared molecular sequence data from 3 genes and a variable region of the poxvirus genome (Table 1) among BRZ-VACV isolates, available vaccine strains related to those used during the eradication campaign in Brazil, and other VACVs isolated from domestic animals (including endemic buffalopox virus in India [15–17] and horsepox virus [HSPV] from Asia [18]).
Sequences were collected from GenBank (Tables 1 and 2). Some isolates had limited sequence data for the 4 genes examined. Although all 4 genes are not available from Lister (B19R gene is not present in the Lister genome), the Lister isolate was included because of its importance in the history of vaccination in Brazil (3,6). Sequences from 3 genes were manually aligned and trimmed to include only the regions available for all isolates (VACV-COP A56R 161210–162095, E3L 51465–50929, B19R 178242–179173). Nucleotide identities were calculated by using BioEdit (www.mbio.ncsu.edu/bioedit/bioedit.html) (Technical Appendix). PAUP* version 4 (Sinauer Associates; Sunderland, MA, USA) and MrBayes 3.1 (http://mrbayes.csit.fsu.edu) were used to analyze a matrix of the 3 concatenated genes (19,20). Parsimony analysis was performed on 1,000 random addition replicates. Appropriate models of sequence evolution were determined by Modeltest (21) with the Akaike information criterion (22,23). The identified model was then applied to maximum likelihood and Bayesian analyses. In MrBayes, the Markov chain Monte Carlo searches explored 4 chains for 1 million generations, sampling every 100 generations. Sample points during the first 50,000 generations were discarded as burn-in, before which the chain reached stationarity (fixed condition). To account for any considerable rate variation between genes, a second Bayesian analysis was run wherein each gene was partitioned and allowed to vary independently (MrBayes command: prset ratepr = variable). DNA sequence data for the fourth coding region, rpo132-ATI-p4c-A27 (A-type inclusion body gene), were not included in the phylogenetic analysis; however, published digestion profiles from relevant strains were compared.
Figure 2
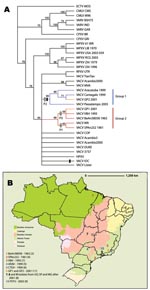
Figure 2. A) Strict consensus of the 6 most parsimonious trees derived from Orthopoxvirus species on the basis of B19R, E3L, and A56R sequences and rooted with ectromelia virus from Moscow (ECTV-MOS). Bootstrap...
The resultant phylogenetic tree (Figure 2A) depicts several highly supported clusters including monophyletic VACV, MPXV, VARV, and camelpox virus (CMLV) assemblages. VARV and CMLV isolates are depicted as sister taxa with high support. Brazilian isolates fall into 2 well-supported monophyletic groups. Group 1 comprises Araçatuba virus (ARAV), Cantagalo virus (CTGV), Guarani P2 virus (GP2V), and Passatempo virus (PSTV); group 2 comprises GP1V, Belo Horizonte virus (VBH), BeAn 58058 virus (BAV), SPAn232 virus (SAV), and VACV-Western Reserve (WR). Results of all analyses—parsimony, maximum likelihood, and Bayesian analysis (data not shown)—all generated 2 distinct Brazilian clades and indicated a close relationship between group 2 and VACV-WR. Neither group of BRZ-VACV was directly linked with VACV-Oswaldo Cruz Institute (IOC) or VACV-Lister in any analysis. Both BRZ-VACV groups are represented by 4 isolates each. VACV-Acambis3000 and VACV-modified vaccinia Ankara (MVA) group together with high support (98, Figure 2A), which reflects their derivation from vaccinia isolate Ankara (both are virus isolates of MVA). Isolates with known origins from Dryvax (Wyeth Laboratories, Marietta, PA, USA) (VACV-3737, -Acambis3, -Acambis2000, -DUKE) did not group together consistently. This could be a result of numerous passages, the history of Dryvax as a nonclonal vaccine strain, or both.
Genetic comparisons of the analyzed VACV strains can be seen in the Technical Appendix. For BRZ-VACV at all 3 loci, the highest identity values occurred within each phylogenetic group. The E3L sequence of PSTV is the only exception. The PSTV E3L sequence is identical to those of GP1 and WR and showed the next highest identity level with the VBH sequence. This finding may indicate recombination in this portion of the genome between isolates from these 2 groups. Additional genomic data and analysis are needed.
Figure 3
Figure 3. A) Partial amino acid alignment of A56R vaccinia virus Copenhagen (VACV-COP). Sequences were retrieved from GenBank and aligned using the BioEdit program (www.mbio.ncsu.edu/bioedit/bioedit.html). VACV-COP (M35027, 161908-161957) is used as the...
The sequence diversity of the orthopoxvirus hemagglutinin (HA) gene has made it a potential marker for molecular diagnostics and phylogenetics (24). In particular, an 18-nt deletion within the gene has been proposed as an identifier of BRZ-VACV strains (8,11,25). A recent article describes 2 approaches for identifying BRZ-VACV–like isolates (25), both of which rely on presence of the 18-nt deletion. Figure 3 shows that while some Brazilian strains (ARAV, CTGV, GP2V, PSTV) share this deletion, others (SAV, BAV, GP1V and VBH) do not and would not be detected by these methods. Within the variably deleted region (Figure 3, amino acids 245–255), VACV isolates can be grouped into 4 types; Brazilian isolates fall into 2 of these. Rabbitpox virus from Utrecht has a unique deletion of only 2 residues (254, 255); VACV-3737 and VACV-Malbran share a 5–amino acid deletion (245–249). The VACV-3737 isolate was “plaque-purified from a vaccinia lesion following vaccination with Dryvax” (GenBank,accession no. DQ377945); VACV Malbran was used in vaccination programs in Argentina between 1937 and 1970 (27). GP1V and VBH share the same amino acid sequence (ADLYDTYNDND; 245–255), identical to that of VACV-WR, VACV-Lister, and several other strains. The remaining isolates from Brazil share the familiar deletion of 6 amino acids (250–255) described previously. This deletion is also present in the VACV-IOC vaccine isolate (absent in Lister and VACV-WR). The variation in this region demonstrates that the deletion itself is not representative of all Brazilian isolates and therefore is not useful as an identifier of Brazilian VACV.
This same deletion has been recognized as a possible shared derived trait between VAVC-IOC and BRZ-VACV. Although this scenario is indeed possible for the CTGV-like BRZ-VACV of group 1, those isolates of group 2 are unlikely to be derived from the IOC vaccine strain because this strain does not carry the insertion. Furthermore, another variable gap occurs in this region of HA among isolates derived from VACV-Lister and Dryvax (New York City Board of Health [NYCBH]). Some clones present the deletion; others do not. The sequence can be found directly adjacent upstream of the variable deletion of the group 1 Brazilian isolates (Figure 3). Perhaps this region is nonessential and able to easily withstand such variation without detrimental effects on the utility of the HA protein or the fitness of the virus. Regardless, this deletion clearly cannot be relied upon to identify isolates directly related to Lister or NYCBH strains. A single shared deletion is only 1 character toward determining phylogenetic relatedness among strains. If smallpox vaccines are not clones but rather pools of virions that vary at the molecular level, assuming relatedness based on a single molecular variation is even more difficult. Our phylogenetic analyses of 302 informative characters, including variation across the HA gene, does not indicate a close relationship between VACV-IOC and BRZ-VACV. The consensus tree (Figure 2A) depicts the IOC vaccine strain in an unresolved basal polytomy within the vaccinia cluster and is thus inconclusive as to the relatedness of VACV-IOC and BRZ-VACV.
Finally, published data indicate a large amount of variability in the region that includes the A-type inclusion body gene of the BRZ-VACV isolates (5,9,11–13). Figure 3B illustrates the arrangement of open reading frames in this region (rpo132-ATI-p4c-A27L) for CPXV Brighton Red, VACV Copenhagen, VACV-WR, and BRZ-VACV. The formation of A-type inclusion bodies is restricted to cells infected with CPXV, ectromelia virus, and raccoonpox virus. The C-terminus of the gene encoding the ATI protein is highly conserved among orthopoxviruses because it overlaps with the C-terminus of the RNA polymerase (132) coding region (9,13). However, the region between this conserved sequence and the N-terminus of the P4c precursor shows considerable variation among orthopoxvirus strains (26). Deletions and nucleotide changes generate variable coding regions and distinguishable digest patterns in this portion of the genome. Three profiles of this region have been discerned among BRZ-VACV; 2 are differentiated by distinct digestion patterns (6,7,11,13). The third state is the near complete absence of the A26L gene except for the last 112 nt (9,13). According to da Fonseca et al., SAV, a Brazilian VACV isolate, shows an A26L digestion profile identical to that of the VACV-WR strain (profile 1) (6). ARAV, PSTV, and GP2V show a digestion profile compatible with but not identical to that of the SAV, VACV-WR, and Lister strains (profile 2) (7,11,13). In BAV, VBH, and GP1V, a major portion of the A26L gene is missing (profile 3) (5,9,13). The open reading frame is small and is probably not expressed, although this has not been verified by Northern or Western blot. Damaso and colleagues demonstrated that A26L is present in CTGV; however, the digestion profile has not been established. Western blot analysis detected a 94-kDa protein typical of VACV-WR and VACV-IOC (8). These 3 profiles correspond to 3 clades recovered in the phylogenetic tree (Figure 2A). These findings constitute a major distinction between CTGV (also SAV, ARAV, PSTV, and GP2V, which exhibit profiles 1 and 2) and the other Brazilian isolates BAV, VBH, and GP1V (profile 3). Furthermore, the unusual deletion presented by BAV, VBH, and GP1V is remarkably different from the corresponding regions of VACV-IOC, -WR, and -Lister, strains implicated in the release of VACV in Brazil.
Although eradication had been achieved in Europe by 1953 and in Central and North America by 1951, Brazil did not conduct a nationwide vaccination campaign during the 1950s. However, Brazilian people in cities and towns were vaccinated when outbreaks were reported by local authorities (2,3). Not until 1962 did Brazil, working with the Pan American Health Organization, launch a national campaign against smallpox (3). The intensified global WHO program prompted a renewed national program in Brazil in 1966. Fenner et al. (3) present a sample questionnaire that the smallpox eradication unit circulated in 1967 to all vaccine producers in countries accessible to WHO. In Brazil, this included 4 laboratories in different regions: the IOC, the Butantan Institute in São Paulo, the Institute for Biological Research in Porto Alegre, and an institute in the town of Recife, Pernambuco State (3). Records identify the seed strains used at these institutes to be “Paris,” Lister, and “Lederle” (28). “Paris” appears to be a reference to the original animal vaccine imported from the Chambon Institute in 1887. The Institute for Biological Research in Porto Alegre received from London the Lister strain, which was originally developed at the Lister Institute, England. Recife and Butantan both indicated use of a strain obtained from Wyeth’s Lederle Laboratory in the United States. The NYCBH strain was the source of Lederle and forerunner of Dryvax, the live vaccine maintained in the United States since the 1970s, and the research vaccine strain VACV-WR (3). These records confirm information that “Paris,” Lister, and NYCBH were used in Brazil between 1968 and 1971 (2,3). It has been suggested that VACV-WR was used as a vaccine in Brazil as well; however, such use is now thought to be unlikely (6,7). The strain was derived by serial passage in mice infected intracranially and selected for neural virulence to mimic encephalitis, a rare side effect of the vaccine in humans (29,30). When available vaccine supplies dropped to critical levels in 1970, Brazil requested and received reserve vaccine from Argentina at least once (3). The Malbran strain was used in vaccination programs in Argentina until 1970, after which the Lister strain was adopted following WHO recommendations (2,3,27). Vaccine production at the Butantan Institute converted to the Lister strain late in 1970 (3), and Butantan became the official distributor for Brazil. Therefore, at least 4 strains of vaccine might have been distributed within Brazil during the WHO eradication campaign.
Discovery of Brazilian VACV Isolates
Since the 1960s, orthopoxviruses have been repeatedly isolated in Brazil and identified as VACV by classical immunologic, virologic, and molecular methods (4–13,26). In 1963, BAV was isolated from the blood of a rice rat (Oryzomys sp.) captured near the edge of a deforested area bordered by Amazon rain forest (Figure 2B) (4,5). This virus is among the first orthopoxviruses naturally isolated from a wild rodent in Brazil (4,5). In the 1960s and 1970s, the Brazilian government and the Institute Adolfo Lutz conducted surveillance of arboviral activity in forested areas around the city of São Paulo, in the southeastern region of the country. During that investigation, a poxvirus was repeatedly isolated from sentinel mice and called Cotia virus (31). A sample of Cotia, SAV, was sent to the virus laboratory in Minas Gerais State. The specimen was plaque purified in duplicate, and a vaccinialike virus was isolated (6). Samples of Cotia have been studied elsewhere, and independent characterizations of the virus have been contradictory (32–34). These contradictory findings suggest that the material provided in the original sample(s) contained >1 virus, leading to conflicting reports of behavior and serologic relationships. We continue to refer to this sample as SPAn232 with reference to de Souza Lopes et al. (31) and da Fonseca et al. (6) to distinguish it from other isolates described previously.
In 1998, VBH was isolated from frozen clinical samples collected from mice during a mousepox-like outbreak in the animal facility of the Biological Institute of the University of Minas Gerais State in 1993 (Figure 2B) (9). Exanthematous outbreaks affecting dairy cattle and their handlers were reported in Brazil in 1999. Two new VACV strains, ARAV and CTGV, were isolated from sick cows in distinct geographic locations of the southeast region of the country (Figure 2C) (7,8). Since then, an increasing number of similar zoonotic outbreaks have been reported in different countryside areas of Brazil, particularly the southeast and southwest (Figure 2A, B) (10,12). In 2001, GP1V and GP2V were isolated from infected cows from adjacent farms near the area where CTGV had been found. Despite their coincident locale, the 2 new isolates constitute distinctly different VACV isolates (Figure 2A, C) (13). In 2003, PSTV was also isolated from cows in Minas Gerais State during a subsequent bovine vaccinia outbreak (Figure 2C; Table 3 [11]). Isolates from group 1, similar to CTGV, have been collected more often than those from group 2 (10,25), which could reflect greater fitness, prevalence, or virulence associated with group 1; however, any explanation is speculative. The difference might eventually be attributed to an as-yet unidentified sampling bias.
Isolates have been obtained from 3 of 6 Brazilian biome types (Figure 2B). Most have been collected in the Atlantic Forest region, where much of the forest has been cleared for dairy farms and coffee and sugar cane plantations (35). Reported human cases are typically in dairy workers, but SAV was isolated from a sentinel mouse placed within the Cotia Forest, São Paulo State (6), and BAV was obtained from a wild-caught rodent (Oryzomys) in northern Brazil. These occurrences, combined with the genetically similar isolates from the southern portion of the country (2,500 km away), indicate the potential circulation of viral strains throughout Brazil and perhaps other regions of South America. Moreover, some of these genetically divergent strains have a sympatric distribution, as illustrated by the isolation of GP1V (group 2) and GP2V (group 1) from adjacent farms (13). Sequencing of the more rapidly evolving inverted terminal repeats of the genome would help clarify the relationship between BRZ-VACV and its hosts. The near-terminal regions of the genome contain most genes involved in host interaction (14) and may illuminate ways in which the host may have shaped the virus evolution. No data are available regarding host variation, epidemiology, or evidence of clinical manifestations that can distinguish between the 2 genetic BRZ-VACV groups depicted in the phylogenetic analyses.
Potential Origins of Brazilian VACV
The possibility that the BRZ-VACV isolates originated from the spread of a smallpox vaccine strain, particularly VACV-IOC and VACV-Lister, has been proposed (4,6,8). A deletion in HA supports this hypothesis; but the larger portion of evidence, the phylogenetic analysis, does not. Available data offer no solid support for these proposed sources as the origin of BRZ-VACV. The VACV-IOC vaccine should be further examined to determine whether it contains virions without the 18-nt HA deletion. Historical records raise the possibility that strains currently circulating in Brazil were established from the original (first known) introduction of VACV to the region by the slave trade in the early part of the 19th century, by the introduction of the animal-cultivated vaccine in the latter part of the 19th century, or subsequently by the wave of VACV introduction brought through the smallpox eradication program in the mid 20th century. If BRZ-VACV was introduced from the Old World, multiple introductions from essentially the same source region (Europe) over ≈167 years (from at least 1804 to 1971, when vaccination in Brazil ceased) (2,3,36) could be responsible for the observed diversity of BRZ-VACV. Because the source of the naturally occurring isolates of Brazil may well be the same as that of the vaccine strains, the history of this viral species will be difficult to tease apart without direct sampling of naturally occurring isolates from Europe, particularly England, Germany, and France, where vaccine strains used worldwide are thought to have originated.
Vaccine escape has been hypothesized to account for other VACV isolated from domestic animals, including endemic buffalopox in India and HSPV (MNR-76) in Mongolia (15–18). A limited comparison between the Brazilian sequences and HSPV did not produce a monophyletic group, which makes it unlikely that they are derived from a very recent common ancestor such as one of the common vaccine strains developed in the past century. HSPV currently has the only complete genome available for comparison from a naturally occurring isolate. As the initial phylogenetic analysis of this sample suggests, we may find potential sources, such as group 1 of the BRZ-VACV, for vaccine strains circulating in various parts of the globe. Better sampling of naturally occurring VACV will be essential for determining the existence of indigenous VACV in Brazil.
The existence of 2 distinct groups of BRZ-VACV is clear. Their origins and how they are related remain undetermined. When more data become available, the 2 Brazilian groups may be found to be more closely related than illustrated by the tree in Figure 2. No epidemiologic data or clinical manifestations that differentiate isolates from the 2 groups have been reported. Epidemiologic data from recent and current zoonotic outbreaks could help elucidate what the genomic diversity implies. The data do establish a clear connection between group 2 (SAV, BAV, GP1V, and VBH) and NYCBH. VACV-WR is strictly a laboratory strain and has never been used as a vaccine (29,30). Complete genome sequencing of BRZ-VACV isolates and a sampling of clones from the original seed sample of NYCBH would be a dramatic step toward discerning the relationship between them. What differentiates the 2 lineages—history, ecology or recognizable phenotype—should be further investigated. The circulation of multiple variants across overlapping regions raises the possibility of recombination among variants, which may complicate the evolutionary history of BRZ-VACV.
The certification of global eradication of smallpox was an unprecedented event in human history. However, even now, the origins of VACV in nature and as a vaccine remain a mystery (3). During the smallpox eradication campaign, the dogma held that vaccine strains could not survive in nature and that wild-type vaccinia virus was extinct, yet VACV clearly persist today in Brazil and other parts of the world. If these isolates constitute a recently established zoonotic disease, they present a unique opportunity for understanding how molecular changes, such as recombination or other mutations, allow for the adaptation of poxviruses to new hosts and ecologic pressures.
Dr Trindade is a virologist and a guest researcher at the Centers for Disease Control and Prevention in Atlanta, Georgia, USA. She has been actively involved in research projects examining the emergence of Vaccinia virus in Brazil. Her areas of interest include emergent viruses, molecular biology and viral diversity.
Acknowledgment
The authors are grateful to Russell Regnery and Mary Reynolds for critical reviews of earlier versions of this manuscript.
References
- Lewis-Jones S. Zoonotic poxvirus infections in humans.Curr Opin Infect Dis. 2004;17:81–9. DOIPubMedGoogle Scholar
- Fernandes T. The smallpox vaccine: its first century in Brazil (from the Jennerian to the animal vaccine)[in Spanish]. Hist Cienc Saude Manguinhos. 1999;6:29–51. DOIPubMedGoogle Scholar
- Fenner F, Henderson DA, Arita I, Jezek Z, Ladnyi ID. Smallpox and its eradication. Geneva: World Health Organization; 1988.
- Fonseca FG, Lanna MC, Campos MAS, Kitajima EW, Peres JN, Golgher RR, Morphological and molecular characterization of the poxvirus BeAn 58058.Arch Virol. 1998;143:1171–86. DOIPubMedGoogle Scholar
- Marques JT, Trindade GS, da Fonseca FG, dos Santos JR, Bonjardim CA, Ferreira PCP, Characterization of ATI, TK and IFN-alpha/betaR genes in the genome of the BeAn 58058 virus, a naturally attenuated wild orthopoxvirus.Virus Genes. 2001;23:291–301. DOIPubMedGoogle Scholar
- da Fonseca FG, Trindade GS, Silva RL, Bonjardim CA, Ferreira PC, Kroon EG. Characterization of a vaccinia-like virus isolated in a Brazilian forest.J Gen Virol. 2002;83:223–8.PubMedGoogle Scholar
- de Souza Trindade G, da Fonseca FG, Marques JT, Nogueira ML, Mendes LC, Borges AS, Araçatuba virus: a vaccinialike virus associated with infection in humans and cattle.Emerg Infect Dis. 2003;9:155–60.PubMedGoogle Scholar
- Damaso CR, Esposito JJ, Condit RC, Moussatche N. An emergent poxvirus from humans and cattle in Rio de Janeiro State: Cantagalo virus may derive from Brazilian smallpox vaccine.Virology. 2000;277:439–49. DOIPubMedGoogle Scholar
- Trindade GS, da Fonseca FG, Marques JT, Diniz S, Leite JA, De Bodt S, Belo Horizonte virus: a vaccinia-like virus lacking the A-type inclusion body gene isolated from infected mice.J Gen Virol. 2004;85:2015–21. DOIPubMedGoogle Scholar
- Nagasse-Sugahara TK, Kisielius JJ, Ueda-Ito M, Curti SP, Figueiredo CA, Cruz AS, Human vaccinia-like virus outbreaks in São Paulo and Goias States, Brazil: virus detection, isolation and identification.Rev Inst Med Trop Sao Paulo. 2004;46:315–22. DOIPubMedGoogle Scholar
- Leite JA, Drumond BP, Trindade G, Lobato ZIP, da Fonseca FG, Santos JR, Passatempo virus, a vaccinia virus strain, Brazil.Emerg Infect Dis. 2005;11:1935–8.PubMedGoogle Scholar
- Lobato ZIP, Trindade GS, Frois MCM, Ribeiro EBT, Dias GRC, Teixeira BM, Outbreak of exanthemal disease caused by vaccinia virus in human and cattle in Zona da Mata region, Minas Gerais[in Portuguese]. Arq Bras Med Vet Zootec.2005;57:423–9. DOIGoogle Scholar
- Trindade GS, Lobato ZIP, Drumond BP, Leite JA, Trigueiro RC, Guedes MIMC, Isolation of two vaccinia virus strains from a single bovine vaccinia outbreak in rural area from Brazil: implications on the emergence of zoonotic orthopoxviruses.Am J Trop Med Hyg. 2006;75:486–90.PubMedGoogle Scholar
- Johnson GP, Goebel SJ, Paoletti E. An update on the vaccinia virus genome.Virology. 1993;196:381–401. DOIPubMedGoogle Scholar
- Dumbell K, Richardson M. Virological investigations of specimens from buffaloes affected by buffalopox in Maharashtra State, India between 1985 and 1987.Arch Virol. 1993;128:257–67. DOIPubMedGoogle Scholar
- Kolhapure RM, Tupe CD, Deolankar RP, Raut CG, Basu A, Dama BM, Investigation of buffalopox outbreaks in Maharashtra State during 1992-1996.Indian J Med Res. 1997;106:441–6.PubMedGoogle Scholar
- Singh RK, Hosamani M, Balamurugan V, Satheesh CC, Rasool TJ, Yadav MP. Comparative sequence analysis of envelope protein genes of Indian buffalopox virus isolates.Arch Virol. 2006;151:1995–2005. DOIPubMedGoogle Scholar
- Tulman ER, Delhon G, Afonso CL, Lu Z, Zsak L, Sandybaev NT, Genome of horsepox virus.J Virol. 2006;80:9244–58. DOIPubMedGoogle Scholar
- Swofford DL. PAUP*. Phylogenetic Analysis Using Parsimony (*and other methods). Version 4. Sunderland (MA): Sinauer Associates; 2003.
- Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees.Bioinformatics. 2001;17:754–5. DOIPubMedGoogle Scholar
- Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models.Bioinformatics. 2003;19:1572–4. DOIPubMedGoogle Scholar
- Posada D, Crandall KA. MODELTEST: testing the model of DNA substitution.Bioinformatics. 1998;14:817–8. DOIPubMedGoogle Scholar
- Posada D, Buckley TR. Model selection and model averaging in phylogenetics: advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests.Syst Biol. 2004;53:793–808. DOIPubMedGoogle Scholar
- Ropp SL, Jin Q, Knight JC, Massung RF, Esposito JJ. PCR strategy for identification and differentiation of smallpox and other orthopoxviruses.J Clin Microbiol. 1995;33:2069–76.PubMedGoogle Scholar
- Damaso CR, Reis SA, Jesus DM, Lima PS, Moussatche N. A PCR-based assay for detection of emerging vaccinia-like viruses isolated in Brazil.Diagn Microbiol Infect Dis. 2006;57:39–46. DOIPubMedGoogle Scholar
- Meyer H, Damon IK, Esposito JJ. Orthopoxvirus diagnostics. In: Isaacs, SN, editor. Vaccinia virus and poxvirology: methods and protocols. Totowa (NJ): Humana Press; 2004 p. 119–33.
- Lewis A, Bok K, Perez O, De Fillipo J, Paolazzi C, Gomes JA. Characterization of a Vaccinia virus strain used to produce smallpox vaccine in Argentina between 1937 and 1970.Arch Virol. 2005;150:1485–91. DOIPubMedGoogle Scholar
- World Health Organization. Archives of the Smallpox Eradication Programme. [cited 2007 May 17.] Available from http://www.who.int/archives/fonds_collections/bytitle/fonds_6/en/index.html
- Turner GS. Respiratory infection of mice with vaccinia virus.J Gen Virol. 1967;1:399–402. DOIPubMedGoogle Scholar
- Wokatsch R. Vaccinia virus. In: Majer M, Plotkin SA, editors. Strains of human viruses. Basel: S. Karger; 1972.
- de Souza Lopes O, Lacerda JP, Fonseca IE, Castro DP, Forattini OP, Rabello EX. Cotia virus: a new agent isolated from sentinel mice in São Paulo, Brazil.Am J Trop Med Hyg. 1965;14:156–7.PubMedGoogle Scholar
- Ueda Y, Dumbell KR, Tsuruhara T, Tagaya I. Studies on Cotia virus—an unclassified poxvirus.J Gen Virol. 1978;40:263–76. DOIPubMedGoogle Scholar
- Ueda Y, Morikawa S, Watanabe T. Unclassified poxvirus: characterization and physical mapping of Cotia virus DNA and location of a sequence capable of encoding a thymidine kinase.Virology. 1995;210:67–72. DOIPubMedGoogle Scholar
- Esposito JJ, Palmer EL, Borden EC, Harrison AK, Obijeski JF, Murphy FA. Studies on the poxvirus Cotia.J Gen Virol. 1980;47:37–46. DOIPubMedGoogle Scholar
- Conservational International [cited 2006 Sep 2]. Available from http://www.biodiversityhotspots.org/xp/Hotspots/atlantic_forest
- Benchimol JL. Febre Amarela: a doença e a vacina, uma historia inacabada. Rio de Janeiro: Editora Fiocruz; 2001.
Figures
Tables
Cite This ArticleTable of Contents – Volume 13, Number 7—July 2007
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Inger Damon, Centers for Disease Control and Prevention, 1600 Clifton Rd NE, Mailstop G43, Atlanta, GA 30333, USA;
Top