Volume 22, Number 2—February 2016
Dispatch
Bordetella pertussis Strain Lacking Pertactin and Pertussis Toxin
Abstract
A Bordetella pertussis strain lacking 2 acellular vaccine immunogens, pertussis toxin and pertactin, was isolated from an unvaccinated infant in New York State in 2013. Comparison with a French strain that was pertussis toxin–deficient, pertactin wild-type showed that the strains carry the same 28-kb deletion in similar genomes.
Pertussis has resurged in the United States in recent decades; >48,000 cases were reported in 2012 (http://www.cdc.gov/pertussis/surv-reporting/cases-by-year.html). Suggested causes include improved surveillance and diagnostics, waning immune response to acellular vaccines introduced in the United States in the 1990s (DTaP [diphtheria, tetanus, and pertussis]; Tdap, [tetanus, diphtheria, and pertussis]), and changes to circulating B. pertussis strains, which led to a mismatch with vaccine strains (1). Components of acellular pertussis vaccines in the United States are pertactin (Prn), pertussis-toxin (Pt), filamentous hemagglutinin, and sometimes fimbrial proteins 2/3. Since 2010, multiple mutations have been documented in the Prn-encoding gene (prn), which have spread rapidly across the United States and other countries (2,3). Pt-deficient Bordetella pertussis isolates are rare, with 1 report from France (4). To our knowledge, B. pertussis that lacks Pt and an additional acellular vaccine immunogen has not been documented.
Prodromal pertussis symptoms developed on March 4, 2013, in an 11-month-old white, non-Hispanic infant from New York State while the family was traveling outside the state. Cough reportedly began on March 14, 2013, and 12 days later (March 26) he was brought to his healthcare provider (HCP) with symptoms consistent with pertussis. Since the child’s birth, the diagnosing HCP had seen the child only once; no visits to other HCPs were known. Per parental report, the case-patient was experiencing paroxysmal cough, apnea, and posttussive vomiting. No thoracic radiograph was obtained. A 5-day course of oral azithromycin was prescribed; the parent reported that the infant received treatment for 3 consecutive days, beginning March 26, 2013. The infant was not reported to have any pertussis-associated complications (seizures, pneumonia, or encephalopathy) and had only light coughing as of April 11, 2013.
The infant was unvaccinated because the parents refused administration of all vaccines. Three siblings, ages 12, 10, and 8 years, lived with the infant and were undervaccinated; they had received 2, 1, and 3 doses, respectively, of pertussis-containing vaccines. No coughing illness was reported among the siblings. The mother reported that she received Tdap vaccine during her pregnancy with the case-patient, but receipt of vaccine could not be verified.
A nasopharyngeal swab specimen was collected from the infant on March 26, 2013, for testing at a commercial laboratory. The isolate was also forwarded to New York State’s public health laboratory, the Wadsworth Center, where it was found to be positive for B. pertussis by PCR targeting IS481 and BP283 (5). Both laboratories yielded positive culture results for B. pertussis. No other testing was performed.
The Wadsworth Center forwarded the isolate, designated I979, to the Centers for Disease Control and Prevention (CDC; Atlanta, Georgia, USA) for confirmatory identification and molecular typing as part of the Enhanced Pertussis Surveillance program (6). PCR amplification of the gene encoding the first subunit of Pt (ptxA) was unsuccessful while the CDC multitarget real-time PCR diagnostic assay was performed (7). Amplification of the promoter region (ptxP) and ptxA was also unsuccessful during multilocus sequence typing targeting acellular vaccine component genes ptxA, ptxP, prn, and fim3 (8).
Further characterization of I979 and French strain FR3749 (4) was undertaken by multilocus sequence typing, multilocus variable-number tandem-repeat analysis, pulsed-field gel electrophoresis (9), and whole-genome sequencing. Long sequencing reads were obtained with the Pacific Biosciences RS II (Menlo Park, CA, USA) at >120× coverage and assembled de novo into a single contig by using HGAP v3 and Quiver v1 (Pacific Biosciences). Assembly structure was confirmed with a genome optical map after restriction digestion with KpnI (OpGen, Gaithersburg, MD, USA). The final sequence was polished with short reads obtained with Illumina MiSeq and CLC Genomics Workbench v7.5.1 (QIAGEN, Valencia, CA, USA) with >90× coverage. Completed genomes were submitted to the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/) with GenBank accession nos. CP010965 (FR3749) and CP010966 (I979). Basic genome metrics are listed in Table 1.
Figure 1
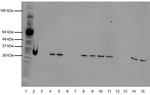
Figure 1. Western blot of pertussis toxin (Pt) expression in Bordetella pertussis Tohama I, I979, FR3749, and 3 additional recent isolates. All isolate lanes were loaded with 10-μg of protein, extracted after growth...
Prn production was determined by ELISA (2,11). Pt production was examined through Western blot analysis of cultures grown in cyclodextrin-modified Stainer-Scholte liquid medium to optical density (OD) 600 nm = 0.1 (12). Proteins precipitated with trichloroacetic acid were washed, reduced, and separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Pt detection is described in Figure 1.
I979 and FR3749 share the same multilocus variable-number tandem-repeat analysis type 27 and prn-2 genotype, the most common recent types (3,13). I979 and FR3749 are fim3–1 and fim3–2, respectively. The fim3 locus has fluctuated between these 2 alleles recently (3,8). Pulsed-field gel electrophoresis indicated that I979 displays profile CDC306, and FR3749 displays CDC046. I979 and FR3749 both lacked Pt production, as shown by Western blot (Figure 1). I979 also failed to produce Prn, whereas FR3749 was positive for Prn production by ELISA, within the range of negative (OD 0.3–0.6) and positive (OD 1.2–1.6) controls.
Figure 2

Figure 2. Map of the 28-kb region deleted in Bordetella pertussis strains I979 and FR3749, compared with vaccine strain Tohama I. Vertical lines indicate the deletion boundaries, at a nucleotide G of the...
Comparison of assembled I979 and FR3749 genomes with that of Tohama I (GenBank accession no. NC_002929.2) (10) indicated that the entire ptx/ptl operon is missing as the result of a putative deletion spanning 28,040 bp (Figure 2). Both genomes contain a conserved, truncated IS481 immediately upstream of the deletion and a single IS481 (FR3749) or 2 tandem IS481 sequences (I979) immediately downstream (Figure 2). Within Tohama I, the region absent from I979 and FR3749 encodes 30 predicted genes bound by 2 NCATGN motifs, the target sequence for IS481 insertion (Table 2). The insertion element IS1002 is located within the 3′ end of this region, and shares a GCATGG motif with IS481 immediately downstream. The 3′ deletion boundary is between IS1002 and IS481 (Figure 2). Whole-genome alignment, using progressiveMauve (14), of I979 and FR3749 with Tohama I revealed structural variation through genomic rearrangements and inversions. In particular, I979 and FR3749 genomes differ by a single, large inversion, the coordinates of which correspond to 2 conserved insertions of IS481 in opposing orientations (Technical Appendix Figure). I979 and FR3749 differ by 31 single nucleotide polymorphisms, each differing from Tohama I by 204 and 173 single nucleotide polymorphisms, respectively. FR3749 contains wild-type prn at position 1613, whereas I979 prn contains an IS481 insertion, the most common cause of Prn-deficiency, at position 1613 (2,11).
B. pertussis strain I979, identified in our study, is both Prn- and Pt-deficient. Loss of Pt in B. pertussis is a rare occurrence; only 2 isolates have been documented in 8 years. Both I979 and FR3749 were isolated from unvaccinated infants (11 months and 3 months old, respectively), who exhibited typical pertussis symptoms, although FR3749 had difficulty colonizing and multiplying in respiratory tracts of adult mice (4). B. pertussis isolates with deletions at other sites across the genome, including part or all of prn, were reported previously (4,15). During the past 5 years, US B. pertussis isolates have become nearly 100% Prn-deficient (2,3) (unpub. data), and Prn-deficient isolates have been obtained from vaccinated persons (11). The loss of Pt may represent a higher fitness cost to B. pertussis than the loss of Prn. In addition, the possibility that only the Pt-deficient isolates were recovered from patients who were co-infected with wild-type and mutant B. pertussis cannot be discarded. Further testing in models to understand the clinical relevance of Prn- and Pt-deficient strains in vaccinated and unvaccinated persons is warranted.
Although incidence of combined Pt- and Prn-deficiency in B. pertussis is rare, any increased mutation in these or other acellular vaccine immunogens may have serious implications for the efficacy of current vaccines. Global epidemiologic, culture-based, and molecular-based monitoring of B. pertussis is critical for understanding current trends of the disease it causes.
Dr. Williams is a research scientist in the Pertussis and Diphtheria Laboratory, Division of Bacterial Diseases, National Center for Immunization and Respiratory Diseases, CDC, Atlanta, Georgia, USA. She specializes in molecular characterization of bacterial pathogens, with a current focus on pertussis genomics.
Acknowledgments
B. pertussis isolate I979 was contributed by the state of New York through participation in the Enhanced Pertussis Surveillance of the Emerging Infections Program Network. In particular, we thank Kathy Root, Kimberlee Musser, and Tammy Quinlan for providing clinical data and isolate I979 and for performing pertussis PCR testing. We also thank Nicole Guiso, Institut Pasteur, Paris, France, for sharing isolate FR3749.
Funding for genome sequencing was provided internally by CDC as part of its Advanced Molecular Detection Initiative.
Members of the CDC Pertussis Working Group: Dhwani Batra, Kanwar Bedi, Katherine E. Bowden, Pamela K. Cassiday, Marsenia J. Harrison, Owen Herzegh, Kristen Knipe, Vladimir Loparev, Lucia C. Pawloski, Yanhui Peng, Lori A. Rowe, Scott Sammons, Mili Sheth.
References
- Misegades LK, Winter K, Harriman K, Talarico J, Messonnier NE, Clark TA, Association of childhood pertussis with receipt of 5 doses of pertussis vaccine by time since last vaccine dose, California, 2010. JAMA. 2012;308:2126–32. DOIPubMedGoogle Scholar
- Pawloski LC, Queenan AM, Cassiday PK, Lynch AS, Harrison MJ, Shang W, Prevalence and molecular characterization of pertactin-deficient Bordetella pertussis in the United States. Clin Vaccine Immunol. 2014;21:119–25 . DOIPubMedGoogle Scholar
- Bowden KE, Williams MM, Cassiday PK, Milton A, Pawloski L, Harrison M, Molecular epidemiology of the pertussis epidemic in Washington State in 2012. J Clin Microbiol. 2014;52:3549–57. DOIPubMedGoogle Scholar
- Bouchez V, Brun D, Cantinelli T, Dore G, Njamkepo E, Guiso N. First report and detailed characterization of B. pertussis isolates not expressing pertussis toxin or pertactin. Vaccine. 2009;27:6034–41. DOIPubMedGoogle Scholar
- Probert WS, Ely J, Schrader K, Atwell J, Nossoff A, Kwan S. Identification and evaluation of new target sequences for specific detection of Bordetella pertussis by real-time PCR. J Clin Microbiol. 2008;46:3228–31. DOIPubMedGoogle Scholar
- Skoff TH, Baumbach J, Cieslak PR. Tracking pertussis and evaluating control measures through enhanced pertussis surveillance, Emerging Infections Program, United States. Emerg Infect Dis. 2015;21:1568. DOIPubMedGoogle Scholar
- Tatti KM, Sparks KN, Boney KO, Tondella ML. Novel multitarget real-time PCR assay for rapid detection of Bordetella species in clinical specimens. J Clin Microbiol. 2011;49:4059–66. DOIPubMedGoogle Scholar
- Schmidtke AJ, Boney KO, Martin SW, Skoff TH, Tondella ML, Tatti KM. Population diversity among Bordetella pertussis isolates, United States, 1935–2009. Emerg Infect Dis. 2012;18:1248–55. DOIPubMedGoogle Scholar
- Hardwick TH, Cassiday P, Weyant RS, Bisgard KM, Sanden GN. Changes in predominance and diversity of genomic subtypes of Bordetella pertussis isolated in the United States, 1935 to 1999. Emerg Infect Dis. 2002;8:44–9 and. DOIPubMedGoogle Scholar
- Parkhill J, Sebaihia M, Preston A, Murphy LD, Thomson N, Harris DE, Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat Genet. 2003;35:32–40. PubMed DOIGoogle Scholar
- Martin SW, Pawloski L, Williams M, Weening K, DeBolt C, Qin X, Pertactin-negative Bordetella pertussis strains: evidence for a possible selective advantage. Clin Infect Dis. 2015;60:223–7. DOIPubMedGoogle Scholar
- Imaizumi A, Suzuki Y, Ono S, Sato H, Sato Y. Effect of heptakis(2,6-O-dimethyl)β-cyclodextrin on the production of pertussis toxin by Bordetella pertussis. Infect Immun. 1983;41:1138–43 .PubMedGoogle Scholar
- Bart MJ, Harris SR, Advani A, Arakawa Y, Bottero D, Bouchez V, Global population structure and evolution of Bordetella pertussis and their relationship with vaccination. MBio. 2014;5:e01074. DOIPubMedGoogle Scholar
- Darling AE, Mau B, Perna NT. progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE. 2010;5:e11147. DOIPubMedGoogle Scholar
- King AJ, van Gorkom T, Pennings JL, van der Heide HG, He Q, Diavatopoulos D, Comparative genomic profiling of Dutch clinical Bordetella pertussis isolates using DNA microarrays: identification of genes absent from epidemic strains. BMC Genomics. 2008;9:311. DOIPubMedGoogle Scholar
Figures
Tables
Cite This Article1Members of the CDC Pertussis Working Group are listed at the end of this article.
Table of Contents – Volume 22, Number 2—February 2016
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Margaret M. Williams, Centers for Disease Control and Prevention, 1600 Clifton Rd, Atlanta, GA 30329-4027, USA
Top